Les récepteurs couplés aux protéines G (RCPG), qui sont présents de la levure à l’homme, représentent la plus grande superfamille de récepteurs dans de nombreux organismes1. Ils jouent un rôle essentiel dans la régulation de presque tous les processus biologiques chez les animaux. Il y a 50 à 200 RCPG dans le génome des arthropodes, ce qui signifie qu’ils représentent la plus grande superfamille de récepteurs membranaires2. Ils sont classés en six classes principales, A-F, en fonction de leur similitude de séquence et de leurs fonctions3. Les RCPG transduisent divers signaux extracellulaires, tels que ceux des hormones, des neuropeptides, des amines biogènes, du glutamate, du proton, des lipoglycoprotéines et des photons4. Les RCPG se couplent aux protéines G hétérotrimères (Gα, Gβ et Gγ) pour transmettre des signaux en aval. Les RCPG couplés aux protéines Gαs ou Gαi/o augmentent ou diminuent, respectivement, les niveaux intracellulaires d’adénosine monophosphate (AMPc) 3′, 5′-cyclique en activant ou en inhibant l’adénylylcyclase. Les RCPGs couplés à Gαq/11 induisent la libération de calcium à partir des réserves de calcium du réticulum endoplasmique en activant la voie phospholipase C (PLC)-inositol-1,4,5-triphosphate (IP3). Les RCPGs couplés à Gα12/13 activent les facteurs d’échange nucléotidiquesde la RhoGTPase 5,6. Les RCPG sont la cible de plus de 50% des médicaments humains et d’un acaricide, l’amitraze4. Comme les RCPG transduisent des signaux aussi divers, ils sont des cibles prometteuses pour le développement de nouveaux pesticides qui perturbent les fonctions physiologiques spécifiques aux invertébrés.
L’objectif de HTS est d’identifier les molécules de frappe qui peuvent moduler les fonctions des récepteurs. HTS implique le développement de tests, la miniaturisation et l’automatisation7. Les RCPG des neuropeptides arthropodes sont impliqués dans la plupart des fonctions physiologiques, telles que le développement, la mue et l’ecdysis, l’excrétion, la mobilisation énergétique et la reproduction4. La plupart des RCPG neuropeptidiques des arthropodes et des métazoaires signalent par la cascade de signalisation calcique 2,6,8,9,10, comme dans le peptide myoinhibiteur et les récepteurs SIFamide de la tique à pattes noires Ixodes scapularis; leurs ligands sont antagonistes dans les essais de motilité de l’intestin postérieur, le SIF provoquant une contraction et la MIP l’inhibant11,12. Un récepteur de type NPY du moustique de la fièvre jaune, Aedes aegypti, régule l’hôte femelle à la recherche13. Comparé à d’autres tests alternatifs de mobilisation du calcium tels que le test de bioluminescence calcique de l’équorine14, le test de fluorescence du calcium est facile à réaliser, ne nécessite pas la transfection d’autres protéines de détection du calcium recombinant et est rentable. Le test de fluorescence calcique produit un signal prolongé par rapport au signal cinétique rapide obtenu dans le test de bioluminescence calcique de l’équorine14,15.
Dans l’exemple ici, le récepteur kinine de la tique de la fièvre bovine, Rhipicephalus microplus, a été exprimé de manière recombinante dans la lignée cellulaire CHO-K1 et utilisé pour le test de fluorescence calcique. Il n’y a qu’un seul gène récepteur de la kinine trouvé dans R. microplus; le récepteur signale par une voie de signalisation dépendante de la protéine Gq et déclenche l’efflux de Ca2+ des réserves de calcium dans l’espace intracellulaire16. Ce processus peut être détecté et quantifié par un fluorophore, qui provoque un signal de fluorescence lors de la liaison des ions calcium (Figure 1).
Le récepteur kinine est un RCPG spécifique aux invertébrés, qui appartient aux récepteurs de type rhodopsine de classe A. La kinine est un ancien neuropeptide de signalisation présent dans Mollusca, Crustacea, Insecta et Acari 4,17,18. Les coléoptères (coléoptères) n’ont pas le système de signalisation des kinines; chez le moustique Aedes aegypti, il n’y a qu’un seul récepteur de kinine qui se lie à trois aedeskinines, tandis que Drosophila melanogaster a un récepteur de kinine avec la drosokinine comme ligandunique 19,20,21. Il n’y a pas de kinines homologues ou de récepteurs de kinines chez les vertébrés. Bien que la fonction exacte de la kinine soit inconnue chez les tiques, les femelles de R. microplus silencieuses de R. microplus soumises au récepteur de la kinine sont réduites de manière significative22. Les kinines sont des peptides pléotropes chez les insectes. Chez Drosophila melanogaster, ils sont impliqués dans les systèmes de régulation nerveux central et périphérique23, la pré-ecdysis 24, l’alimentation25, le métabolisme 26 et les schémas d’activité du sommeil26,27, ainsi que la locomotionlarvaire 28. Les kinines régulent la contraction de l’intestin postérieur, la diurèse et l’alimentation du moustique A. aegypti 29,30,31. Les peptides kinines ont conservé un pentapeptide C-terminal Phe-X1-X2-Trp-Gly-NH2, qui est la séquence minimale requise pour l’activité biologique32. La spécificité des arthropodes, la petite taille du ligand endogène, qui les rend sujets à l’interférence des petites molécules, et les fonctions pléiotropiques chez les insectes font du récepteur kinine une cible prometteuse pour la lutte antiparasitaire4.
Le test de « double addition » (Figure 2) permet l’identification d’agonistes ou d’antagonistes dans le même test HTS15. Il est adapté d’un test de « double addition » couramment utilisé dans l’industrie pharmaceutique pour la découverte de médicaments33. En bref, le premier ajout de médicaments dans la plaque cellulaire permet d’identifier les agonistes potentiels dans la bibliothèque chimique lorsqu’un signal de fluorescence plus élevé est détecté par rapport à l’application du contrôle du solvant. Après 5 min d’incubation avec ces petites molécules, un agoniste connu (peptide kinine) est appliqué sur tous les puits. Les puits qui ont reçu au hasard un antagoniste de la plaque médicamenteuse affichent un signal de fluorescence plus faible lors de l’ajout d’agonistes par rapport aux puits témoins qui ont reçu le solvant lors de la première addition. Ce test permet ensuite d’identifier des agonistes et antagonistes potentiels avec les mêmes cellules. Dans un projet HTS standard, ces molécules frappées seraient validées par des tests dose-réponse et par des tests d’activité biologique supplémentaires, qui ne sont pas présentés ici.
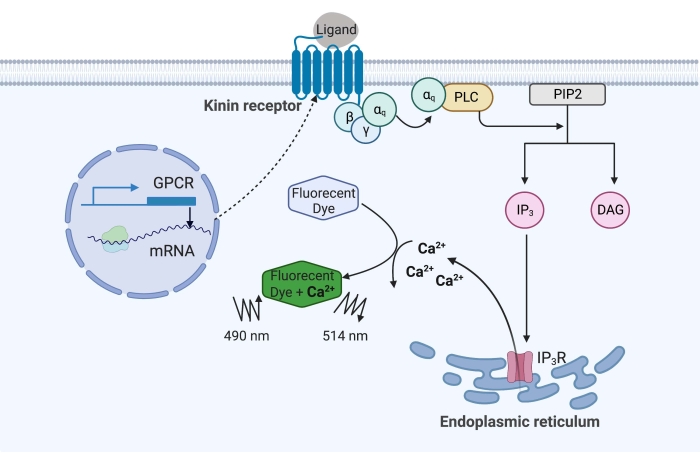
Figure 1 : Illustration du mécanisme d’essai de fluorescence calcique. La protéine Gq déclenche la voie de signalisation intracellulaire du calcium. Le récepteur de la kinine (récepteur couplé à la protéine G) a été exprimé de manière recombinante dans les cellules CHO-K1. Lorsque le ligand agoniste se lie au récepteur, la protéine Gq associée au récepteur kinin active l’API, ce qui catalyse la conversion d’une molécule PIP2 en IP3 et DAG. IP 3 se lie ensuite à l’IP3R à la surface du réticulum endoplasmique, conduisant à la libération de Ca 2+ dans le cytoplasme, où les ions Ca2+ se lient aux fluorophores et provoquent un signal de fluorescence. Le signal de fluorescence peut être obtenu par excitation à 490 nm et détecté à 514 nm. Abréviations : RCPG = récepteur couplé aux protéines G; PLC = phospholipase C; PIP2 = phosphatidylinositol 4,5-bisphosphate; IP3 = inositol trisphosphate; DAG = diacylglycérol; IP3 R = récepteur IP3. Créé avec BioRender.com. Veuillez cliquer ici pour voir une version agrandie de cette figure.
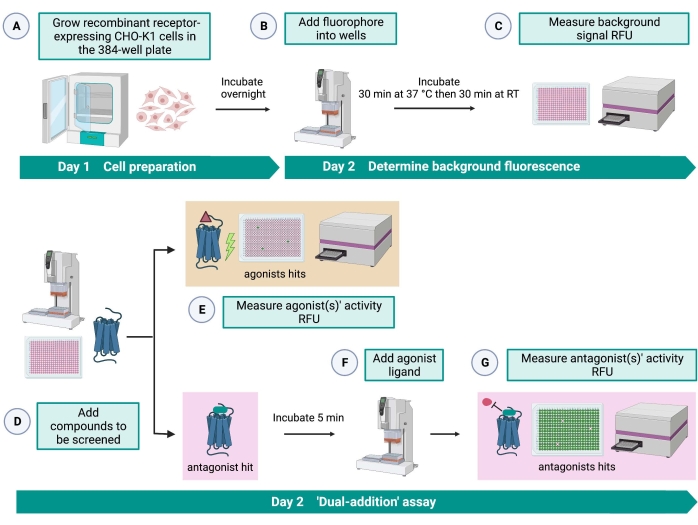
Figure 2 : Flux de travail pour le criblage à haut débit de petites molécules sur un récepteur couplé à une protéine G exprimé dans des cellules CHO-K1. (A) Des cellules CHO-K1 recombinantes exprimant de manière stable le récepteur kinine ont été ajoutées à la plaque de 384 puits (10 000 cellules/puits) à l’aide d’un système de manipulation de liquide (25 μL/puits) et incubées dans un incubateur de CO2 humidifié pendant 12 à 16 h. (B ) Le tampon d’essai contenant le colorant fluorescent (25 μL/puits) a été ajouté dans la plaque cellulaire à l’aide d’un système de manipulation de liquide. La plaque a été incubée pendant 30 min à 37 °C pendant 30 min et équilibrée à TA pendant 30 minutes supplémentaires. (C) Le signal de fluorescence de fond des cellules de chaque puits a été mesuré à l’aide d’un lecteur de plaques. (D) Des solutions médicamenteuses provenant d’une plaque bibliothèque de 384 puits et d’un solvant à blanc (tous à 0,5 μL/puits) ont été ajoutées dans la plaque d’essai cellulaire à l’aide d’un système de manipulation de liquide. (E) Les réponses cellulaires de fluorescence calcique ont été mesurées avec le lecteur de plaque immédiatement après l’ajout des solutions médicamenteuses; Les composés suscitant des signaux de fluorescence supérieurs à la moyenne ont été choisis comme des coups agonistes. Des hits antagonistes qui bloquent le RCPG (icône ci-dessous) ont été révélés après l’ajout de l’agoniste peptidique à l’étape G. (F) Dans la même plaque d’essai, après 5 minutes d’incubation des cellules avec des composés de criblage, un peptide agoniste endogène Rhimi-K-1 (QFSPWGamide) du récepteur des tiques kinines a été ajouté à chaque puits (1 μM). (G) Les réponses de fluorescence cellulaire après l’ajout du peptide agoniste ont été mesurées immédiatement par le lecteur de plaques. Le(s) composé(s) inhibant(s) le signal de fluorescence a été sélectionné comme antagoniste(s). Abréviations : RCPG = récepteur couplé aux protéines G; RT = température ambiante; RFU = unités de fluorescence relative. Créé avec BioRender.com. Veuillez cliquer ici pour voir une version agrandie de cette figure.
