Barriere strette separano il sistema nervoso centrale (SNC) dalla periferia, compresa la barriera emato-encefalica (BBB) e la barriera sangue-liquido cerebrospinale (CSF). Queste barriere proteggono il SNC dagli insulti esterni e garantiscono un microambiente equilibrato e controllato 1,2,3. Mentre la BBB è stata ampiamente studiata nel tempo, la barriera emato-liquorale situata nel plesso coroideo (CP) ha guadagnato solo un crescente interesse di ricerca nell’ultimo decennio. Quest’ultima barriera può essere trovata nei quattro ventricoli del cervello (Figura 1A, B) ed è caratterizzata da un singolo strato di cellule epiteliali del plesso coroideo (CPE) che circondano uno stroma centrale, capillari permeabili, fibroblasti e una popolazione di cellule linfoidi e mieloidi (Figura 1C)4,5,6. Le cellule CPE sono saldamente interconnesse da giunzioni strette, impedendo così la fuoriuscita dai capillari sanguigni fenestrati sottostanti nel liquido cerebrospinale e nel cervello. Inoltre, il trasporto attraverso le cellule CPE è regolato da una serie di sistemi di trasporto verso l’interno e verso l’esterno che gestiscono l’afflusso di composti benefici (ad esempio, nutrienti e ormoni) dal sangue al liquido cerebrospinale e l’efflusso di molecole nocive (ad esempio, rifiuti metabolici, neurotrasmettitori in eccesso) nella direzione opposta 1,6. Per essere in grado di esercitare la loro funzione di trasporto attivo, le cellule CPE contengono numerosi mitocondri nel loro citoplasma7. Inoltre, il CP è la principale fonte di CSF e agisce come il guardiano del cervello dalla presenza di cellule infiammatorie residenti1. Grazie alla sua posizione unica tra il sangue e il cervello, il CP è anche perfettamente posizionato per effettuare la sorveglianza immunitaria8.
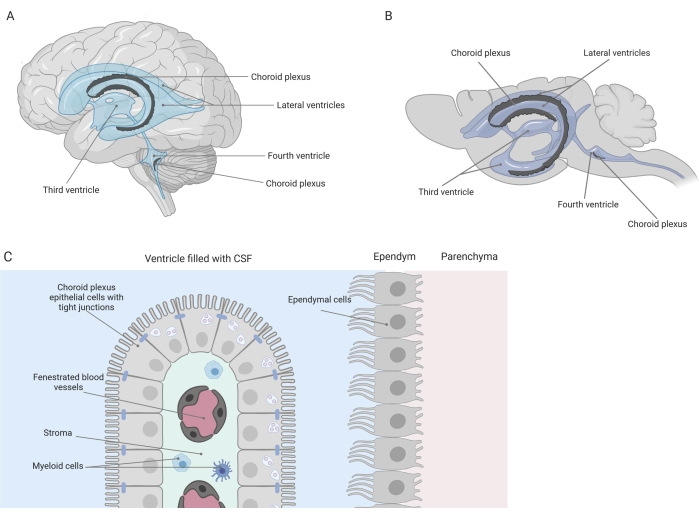
Figura 1: Panoramica schematica della posizione e della composizione del plesso coroideo (CP). (A,B) Il tessuto CP si trova all’interno dei due ventricoli laterali, terzi e quarti del cervello (A) umano e (B) del topo. (C) Il tessuto CP è costituito da un singolo strato di cellule dell’epitelio CP cuboide (CPE) strettamente connesse che circondano i capillari fenestrati, il tessuto connettivo sciolto e le cellule linfoidi e mieloidi, e forma la barriera emato-liquido cerebrospinale (adattata e modificata dal riferimento23). Figura creata con Biorender.com. Fare clic qui per visualizzare una versione ingrandita di questa figura.
Negli ultimi dieci anni, prove crescenti, tra cui diversi rapporti del nostro gruppo di ricerca, hanno rivelato che il CP svolge un ruolo centrale nella salute e nella malattia 9,10,11,12,13,14,15,16,17,18 . Ad esempio, è noto che la barriera emato-CSF che invecchia mostra alterazioni morfologiche, tra gli altri, nei nuclei, nei microvilli e nella membrana basale 1,19. Inoltre, nel contesto della malattia di Alzheimer, l’integrità complessiva della barriera è compromessa e tutti questi cambiamenti legati all’età sembrano essere ancora più pronunciati 1,8,20. Oltre ai cambiamenti morfologici, il trascrittoma, il proteoma e il secretoma della CP sono alterati durante la malattia 12,21,22,23. Pertanto, la conoscenza avanzata della CP è essenziale per comprendere meglio il suo ruolo nelle malattie neurologiche e potenzialmente sviluppare nuove strategie terapeutiche.
Un metodo efficiente per una microdissezione accurata del CP dai ventricoli cerebrali è il primo passo inestimabile per consentire una corretta indagine di questa piccola struttura cerebrale. A causa della sua natura altamente vascolarizzata (Figura 2B), il CP fluttuante all’interno delle cavità ventricolari del cervello può essere identificato utilizzando un microscopio binoculare. Tuttavia, la perfusione transcardica è spesso richiesta per l’analisi a valle, complicando la corretta identificazione e isolamento del tessuto CP (Figura 2C). Se le ulteriori fasi di elaborazione lo consentono (ad esempio, nel caso dell’analisi dell’RNA e delle proteine), la CP può essere visualizzata tramite perfusione transcardica con blu di bromofenolo (Figura 2A). Diverse pubblicazioni descrivono già l’isolamento del CP dal cervello del ratto24 e del topo25. Qui, viene descritta una tecnica di isolamento della microdissezione per isolare il CP dai topi adulti. È importante sottolineare che questa tecnica di isolamento preserva la vitalità, la funzione e la struttura delle cellule all’interno del CP. L’isolamento del CP galleggiante nel quarto ventricolo laterale è descritto qui. In breve, i topi sono anestetizzati terminalmente e, se necessario, perfusi transcardialmente. Tuttavia, va notato che la perfusione può danneggiare la struttura delle cellule all’interno del CP. Di conseguenza, se il campione deve essere analizzato utilizzando la microscopia elettronica a trasmissione (TEM), la microscopia elettronica a scansione a blocchi seriali (SBF-SEM) o il fascio ionico focalizzato SEM (FIB-SEM), la perfusione non deve essere eseguita. Successivamente, l’intero cervello è isolato e il forcipe viene utilizzato per emisecare sagittalmente il cervello. Da qui, i CP che galleggiano nei ventricoli laterali possono essere identificati e sezionati, mentre i CP dal quarto ventricolo possono essere isolati dal lato cerebellare del cervello.
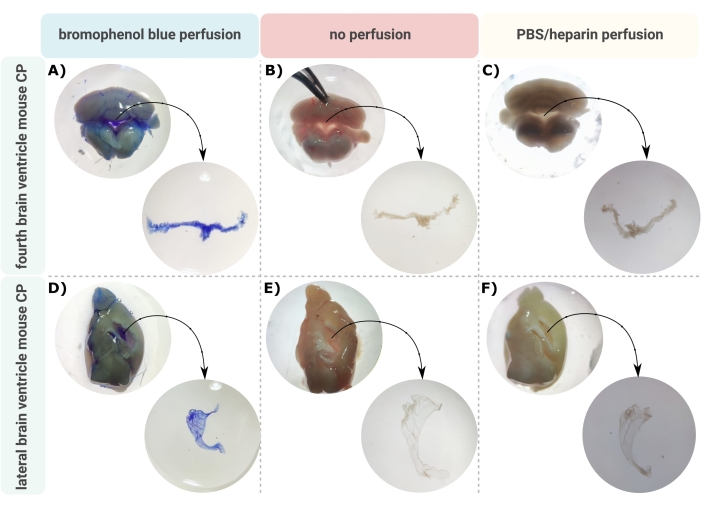
Figura 2: Visualizzazione del quarto plesso coroideo (A-C) e (D-F) ventricolo laterale (CP) dopo (A,D) perfusione blu di bromofenolo, (B,E) nessuna perfusione e (C,F) perfusione con PBS/eparina. Le immagini sono prese con un microscopio stereo (ingrandimento 8x-32x). Fare clic qui per visualizzare una versione ingrandita di questa figura.
Una volta che il CP è correttamente sezionato dai ventricoli cerebrali, un intero repertorio di tecniche può essere applicato per ottenere ulteriori informazioni sulla funzione di questa struttura. Ad esempio, la citometria a flusso o il sequenziamento dell’RNA a singola cellula possono essere eseguiti per quantificare e analizzare fenotipicamente le cellule infiammatorie infiltranti in determinate condizioni di malattia26,27. Oltre alla composizione cellulare, la composizione molecolare del CP può essere analizzata per valutare la presenza di citochine e chemochine tramite saggio immunoassorbente enzimatico (ELISA), immunoblot o attraverso l’analisi simultanea di più citochine utilizzando il citokine bead array28. Inoltre, le analisi del trascrittoma, vascolare, dell’istologia delle cellule immunitarie e del secretoma possono essere eseguite sugli espianti CP microdissecati29. Qui, la microscopia elettronica a scansione (SEM) su CP a montaggio intero viene utilizzata per ottenere una visione d’insieme della struttura CP. SEM utilizza un fascio di elettroni focalizzati per scansionare la superficie e creare un’immagine della topografia e della composizione della superficie. Poiché la lunghezza d’onda degli elettroni è molto più piccola di quella della luce, la risoluzione del SEM è nell’intervallo nanometrico e superiore a quella di un microscopio ottico. Di conseguenza, gli studi morfologici a livello subcellulare possono essere eseguiti tramite SEM. In breve, il CP sezionato viene immediatamente trasferito in un fissativo contenente glutaraldeide per una fissazione notturna, seguita da osmicazione e colorazione dell’acetato di uranile. I campioni vengono quindi trattati con macchie di aspartato di piombo, disidratati e infine incorporati per l’imaging.
Pertanto, questo protocollo facilita l’isolamento efficiente del CP dai ventricoli cerebrali del topo, che può essere ulteriormente analizzato utilizzando una varietà di tecniche a valle per studiarne la struttura e la funzione.
