El cristalino es un tejido transparente y ovoide en la cámara anterior del ojo que está formado por dos tipos de células, epiteliales y fibrosas 1 (Figura 1). Existe una monocapa de células epiteliales que cubre el hemisferio anterior del cristalino. Las células fibrosas se diferencian de las células epiteliales y constituyen la mayor parte del cristalino. Las células fibrosas altamente especializadas experimentan una programación de elongación, diferenciación y maduración, marcada por cambios distintivos en la morfología de la membrana celular desde la periferia del cristalino hasta el centro del cristalino 2,3,4,5,6,7,8,9,10,11,12 , también conocido como núcleo del cristalino. La función del cristalino para enfocar con precisión la luz que proviene de varias distancias sobre la retina depende de sus propiedades biomecánicas, incluida la rigidez y la elasticidad 13,14,15,16,17,18,19. Las complejas interdigitaciones de las fibras del cristalino han sido objeto de la hipótesis 20,21 y recientemente se ha demostrado que son importantes para la rigidez del cristalino 22,23.
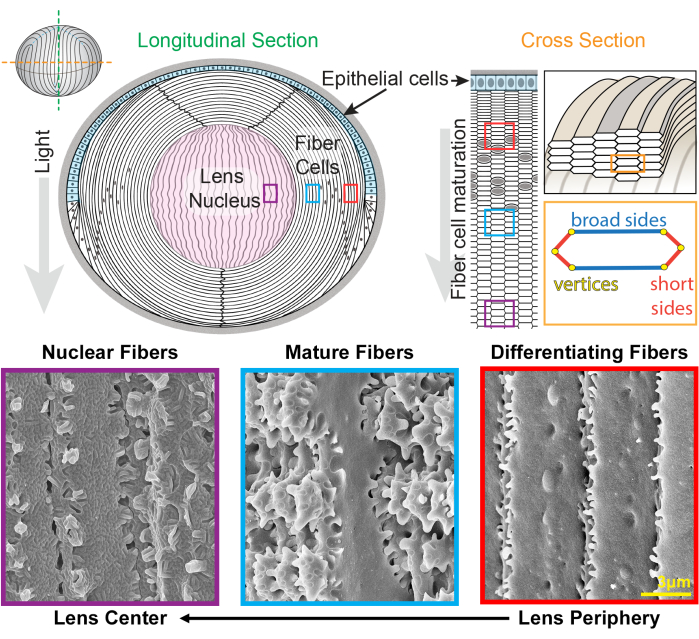
Figura 1: Diagramas de anatomía del cristalino e imágenes representativas de microscopía electrónica de barrido (SEM) de fibras del cristalino. La caricatura muestra una vista longitudinal (anterior a posterior, de arriba a abajo) de la monocapa anterior de células epiteliales (sombreada en azul claro) y una masa masiva de células de fibra del cristalino (blanca). El centro del cristalino (sombreado en rosa) se conoce como núcleo y está formado por células de fibra altamente compactadas. A la derecha, una caricatura de sección transversal revela la forma de celda hexagonal alargada de las fibras de la lente que están empaquetadas en un patrón de panal. Las células de fibra tienen dos lados anchos y cuatro lados cortos. Las imágenes SEM representativas a lo largo de la parte inferior muestran las complejas interdigitaciones de membrana entre las células de fibra de la lente a diferentes profundidades de la lente. Desde la derecha, las fibras recién formadas de la lente en la periferia de la lente tienen pequeñas protuberancias a lo largo de los lados cortos y bolas y zócalos a lo largo del lado ancho (cajas rojas). Durante la maduración, las fibras del cristalino desarrollan grandes dominios de paleta que están decorados por pequeñas protuberancias a lo largo de los lados cortos (cajas azules). Las células fibrosas maduras poseen grandes dominios de paleta ilustrados por pequeñas protuberancias. Estos dominios entrelazados son importantes para las propiedades biomecánicas del cristalino. Las células fibrosas en el núcleo del cristalino tienen menos protuberancias pequeñas a lo largo de sus lados cortos y tienen interdigitaciones machihembradas complejas (cajas moradas). Los lados anchos de la célula muestran una morfología de membrana globular. La caricatura fue modificada a partir de22,32 y no dibujada a escala. Barra de escala = 3 μm. Haga clic aquí para ver una versión más grande de esta figura.
El cristalino crece mediante la adición de capas de nuevas células de fibra superpuestas sobre las generaciones anteriores de fibras24,25. Las células de fibra tienen una forma de sección transversal hexagonal alargada con dos lados anchos y cuatro lados cortos. Estas células se extienden desde el polo anterior hasta el posterior del cristalino y, dependiendo de la especie, las fibras del cristalino pueden tener varios milímetros de longitud. Para apoyar la estructura de estas células alargadas y delgadas, las interdigitaciones especializadas a lo largo de los lados anchos y cortos crean estructuras entrelazadas para mantener la forma de la lente y las propiedades biomecánicas. Los cambios en la forma de la membrana celular durante la diferenciación y maduración de las células de fibra han sido ampliamente documentados por estudios de microscopía electrónica (EM) 2,3,4,5,6,7,8,9,10,20,26,27,28,29 . Las células de fibra recién formadas tienen bolas y alvéolos a lo largo de sus lados anchos con protuberancias muy pequeñas a lo largo de sus lados cortos, mientras que las fibras maduras tienen protuberancias entrelazadas y paletas a lo largo de sus lados cortos. Las fibras nucleares muestran interdigitaciones machihembradas y morfología de membrana globular. Poco se sabe sobre las proteínas que se requieren para estas complejas membranas entrelazadas. Estudios previos sobre la localización de proteínas en células de fibra se han basado en secciones de tejido del cristalino, que no permiten una visualización clara de la compleja arquitectura celular.
Este trabajo ha creado y perfeccionado un método novedoso para fijar células individuales y haces de fibras del cristalino para preservar la morfología compleja y permitir la inmunotinción de proteínas en la membrana celular y dentro del citoplasma. Este método preserva fielmente la arquitectura de la membrana celular, comparable a los datos de los estudios de EM, y permite la tinción con anticuerpos primarios para proteínas específicas. Hemos inmunoteñido previamente fibras corticales del cristalino en proceso de diferenciación y maduración22,23. En este protocolo, también hay un nuevo método para teñir células de fibra del núcleo del cristalino. Este protocolo abre la puerta a la comprensión de los mecanismos de formación y cambios en las interdigitaciones de la membrana durante la maduración de las células de fibra y la compactación del núcleo del cristalino.
