العدسة عبارة عن نسيج شفاف وبيضاوي الشكل في الحجرة الأمامية للعين يتكون من نوعين من الخلايا ، الخلايا الظهارية والليفية 1 (الشكل 1). هناك طبقة أحادية من الخلايا الظهارية التي تغطي نصف الكرة الأمامي للعدسة. تتمايز الخلايا الليفية عن الخلايا الطلائية، وتشكل الجزء الأكبر من العدسة. تخضع خلايا الألياف عالية التخصص لبرمجة الاستطالة والتمايز والنضج ، والتي تتميز بتغيرات واضحة في مورفولوجيا غشاء الخلية من محيط العدسة إلى مركز العدسة2،3،4،5،6،7،8،9،10،11،12، المعروف أيضا باسم نواة العدسة. تعتمد وظيفة العدسة في التركيز الدقيق للضوء القادم من مسافات مختلفة على شبكية العين على خصائصها الميكانيكية الحيوية ، بما في ذلك الصلابة والمرونة13،14،15،16،17،18،19. تم افتراض التداخلات المعقدة لألياف العدسة20,21 وثبت مؤخرا أنها مهمة لصلابةالعدسة 22,23.
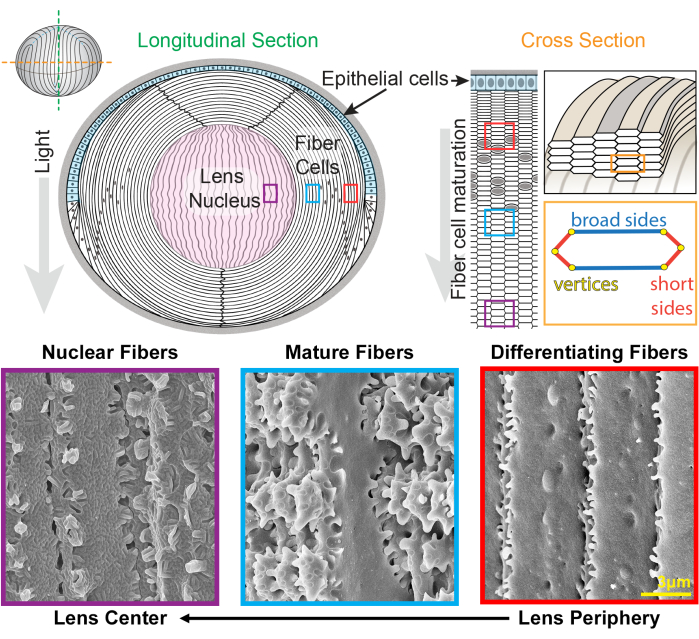
الشكل 1: مخططات تشريح العدسة وصور المجهر الإلكتروني الماسح التمثيلي (SEM) من ألياف العدسة. يظهر الرسم الكاريكاتوري منظرا طوليا (من الأمام إلى الخلف من الأعلى إلى الأسفل) للطبقة الأحادية الأمامية للخلايا الظهارية (مظللة باللون الأزرق الفاتح) وكتلة كبيرة من خلايا ألياف العدسة (بيضاء). يعرف مركز العدسة (المظلل باللون الوردي) باسم النواة ويتكون من خلايا ليفية مضغوطة للغاية. على اليمين ، يكشف رسم كاريكاتوري مقطع عرضي عن شكل الخلية السداسية الممدودة لألياف العدسة المعبأة في نمط قرص العسل. الخلايا الليفية لها جانبان عريضان وأربعة جوانب قصيرة. تظهر صور SEM التمثيلية على طول الجزء السفلي التداخل الغشائي المعقد بين خلايا ألياف العدسة على أعماق مختلفة من العدسة. من اليمين ، تحتوي ألياف العدسة المشكلة حديثا في محيط العدسة على نتوءات صغيرة على طول الجوانب القصيرة والكرات والمقابس على طول الجانب العريض (الصناديق الحمراء). أثناء النضج ، تطور ألياف العدسة مجالات مجداف كبيرة مزينة بنتوءات صغيرة على طول الجوانب القصيرة (الصناديق الزرقاء). تحتوي خلايا الألياف الناضجة على مجالات مجداف كبيرة موضحة بنتوءات صغيرة. هذه المجالات المتشابكة مهمة للخصائص الميكانيكية الحيوية للعدسة. تحتوي الخلايا الليفية في نواة العدسة على عدد أقل من النتوءات الصغيرة على طول جوانبها القصيرة ولها تداخلات معقدة بين اللسان والأخدود (مربعات أرجوانية). تعرض الجوانب العريضة للخلية مورفولوجيا غشاء كروي. تم تعديل الرسوم المتحركة من22,32 ولم يتم رسمها على نطاق واسع. شريط المقياس = 3 ميكرومتر. يرجى النقر هنا لعرض نسخة أكبر من هذا الشكل.
تنمو العدسة عن طريق إضافة أغلفة من خلايا الألياف الجديدة المتراكبة فوق الأجيال السابقة من الألياف24,25. الخلايا الليفية لها شكل مقطع عرضي سداسي ممدود مع جانبين عريضين وأربعة جوانب قصيرة. تمتد هذه الخلايا من القطب الأمامي إلى القطب الخلفي للعدسة ، واعتمادا على الأنواع ، يمكن أن يصل طول ألياف العدسة إلى عدة ملليمترات. لدعم بنية هذه الخلايا الممدودة والنحيفة ، تخلق التداخلات المتخصصة على طول الجانبين العريض والقصير هياكل متشابكة للحفاظ على شكل العدسة والخصائص الميكانيكية الحيوية. تم توثيق التغييرات في شكل غشاء الخلية أثناء تمايز الخلايا الليفية ونضجها على نطاق واسع من خلال دراسات المجهر الإلكتروني (EM)2،3،4،5،6،7،8،9،10،20،26،27،28،29. تحتوي الخلايا الليفية المشكلة حديثا على كرات ومآخذ على طول جوانبها العريضة مع نتوءات صغيرة جدا على طول جوانبها القصيرة ، بينما تحتوي الألياف الناضجة على نتوءات ومجاديف متشابكة على طول جوانبها القصيرة. تعرض الألياف النووية تداخلات اللسان والأخدود ومورفولوجيا الغشاء الكروي. لا يعرف سوى القليل عن البروتينات المطلوبة لهذه الأغشية المتشابكة المعقدة. اعتمدت الدراسات السابقة حول توطين البروتين في خلايا الألياف على أقسام أنسجة العدسة ، والتي لا تسمح بتصور واضح لبنية الخلية المعقدة.
ابتكر هذا العمل وأتقن طريقة جديدة لإصلاح خلايا ألياف العدسة المفردة والحزم للحفاظ على التشكل المعقد والسماح بتلطيخ المناعة للبروتينات في غشاء الخلية وداخل السيتوبلازم. تحافظ هذه الطريقة بأمانة على بنية غشاء الخلية ، مقارنة بالبيانات من دراسات EM ، وتسمح بتلطيخ الأجسام المضادة الأولية لبروتينات معينة. لقد سبق لنا أن خضعنا ألياف العدسات القشرية الملطخة بالمناعة والتي تخضع للتمايز والنضج22,23. في هذا البروتوكول ، هناك أيضا طريقة جديدة لتلطيخ خلايا الألياف من نواة العدسة. يفتح هذا البروتوكول الباب لفهم آليات التكوين والتغيرات في تداخل الغشاء أثناء نضوج الخلايا الليفية وضغط نواة العدسة.
