Development of mouse embryos ex utero depends on multiple factors starting from the time the uterus is isolated from the body to the time the embryos are cultured. As depicted in Figure 1, the procedure involves a series of steps including, separation of the gravid uterus from the body (Figure 1A), isolation of the embryos with intact yolk sac (Figure 1B), exteriorization of the embryos from the yolk sac (Figure 1C) and culturing the embryos in a serum-free media in an atmosphere of 95% O2 / 5% CO2 in a rolling bottle culture apparatus at 37 °C. Embryos exhibited 100% survival when transferred into culture media immediately after separation from the uterus. Under these culture conditions, mouse embryos when cultured for 16-40 hr exhibited growth and morphological development comparable to that observed in equivalent stage embryos developing in utero (Figure 2).
Embryos at E10.5 (~34-35 s) (Figure 2A) when cultured for 16-40 hr survived and advanced in development well beyond the stage of 35 s. The embryos after 16-18 hr in culture (Figures 2D and 3B; Table 1) added about 5-6 s exhibiting a growth rate of one somite for every two and a half hours in culture and were morphologically comparable to E11.0 (~40-41 s) embryos developed in utero (Figures 2B and 3A). Though the development was delayed by about an hour, these embryos in culture exhibited an overall increase in body size and a proportionate head with distinct brain vesicles. The embryos also showed developing limb buds, formation of a well chambered heart and appearance of pigmentation in the retinal pigmented epithelium. However, this type of development was observed in about 58% of the embryos cultured while 28% of the embryos exhibited an intermediate development (Figure 3C). These later embryos showed a smaller body size though they had a similar somite count compared to the in utero developed embryos. They had a relatively smaller head, although the brain vesicles were demarcated. The heart chambers were not distinct and the limb buds in some embryos exhibited dark areas at the extremities. At the other end of the spectrum, about 14% of the embryos exhibited poor development in the culture (Figure 3D). These embryos had shorter body and head sizes and showed dark areas in some parts of the body indicating degenerative changes. Though the heart was beating it was ill developed and lacked differentiation into chambers in some embryos while in others the limbs appeared dark and retarded in growth.
Embryos continued in culture for 38-40 hr (Figure 2E; Table 1) showed about 49-50 s and were comparable to E12.0 embryos (Figure 2C) developed in utero. Although delayed by 2-3 hr, these embryos in culture exhibited proportionate increase in body size and showed appropriate organogenesis and tissue differentiation as observed in the development of the pigmentation in the retinal pigmented epithelium and formation of a well demarcated snout in the head region. Though some parts at the extremities like the limb buds and the tail appeared to be under-developed, the embryos appeared to be developing comparable to the in utero embryos. However, this kind of development was observed in only 30-40% of the embryos cultured (Table 1) while the rest of them exhibited a range of developmental progression with areas of retarded growth or under-development in some parts of the body or the whole embryo.
Apart from the above differences in development, we observed a significant difference in development when the embryos were cocultured in the same culture bottle. Compared to the other embryo (Figures 4A and 4A’) in the coculture, the well-developed embryo (Figures 4B and 4B’) exhibited better growth in all aspects of the embryonic development. Major differences were observed in the overall body size and head size and sometimes in the heart and limb development. While the well-developed embryo from the coculture was morphologically comparable to the embryos developing in utero, the other embryo in the coculture has always appeared to be less developed compared to the well-developed embryo. This kind of difference in development was observed in 45% of the cocultures (N=27), while the rest of the cocultures (N=33) showed almost similar development in both the embryos in the coculture.
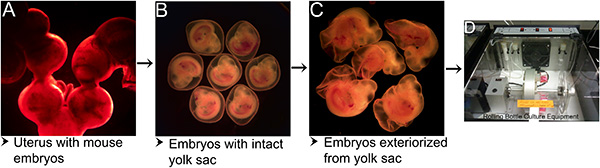
Figure 1. Steps in mouse embryo culture protocol. (A) Gravid uterus with embryos isolated from the mother mouse. (B) Embryos with intact yolk sac separated from decidua after segmental dissection of uterus. (C) Embryos exteriorized from the yolk sac. (D) Rolling bottle culture apparatus at 37 °C and supplied with 95% O2 / 5% CO2 for culturing mouse embryos. Please click here to view a larger version of this figure.
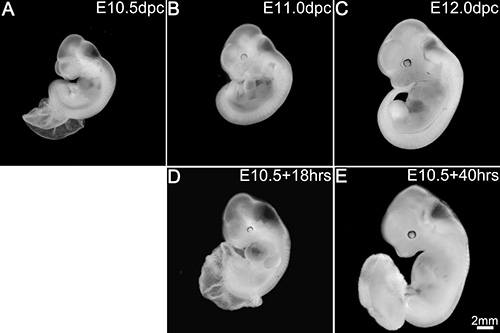
Figure 2. Embryos in culture replicate in utero development. In utero development at E10.5 (A), E11.0 (B) and E12.0 (C). Development in culture after 18 hr (D) and 40 hr (E). Scale bar in (E) applies to all panels. Please click here to view a larger version of this figure.
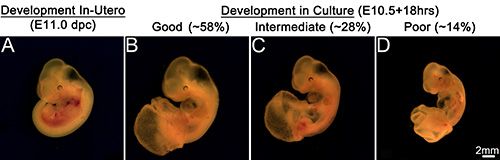
Figure 3. Embryonic development in culture. Embryo developed in utero at E11.0 dpc (A). Development in culture (B, C, D). (B) Representation of good embryonic development in culture. About 58% of total embryos cultured exhibit comparable development to in utero developed embryos. (C) Representation of intermediate development in culture. About 28% of embryos cultured show intermediate development. (D) Representation of poor embryonic development in culture. About 14% embryos show poor development in culture. Scale bar in (D) applies to all panels. Please click here to view a larger version of this figure.
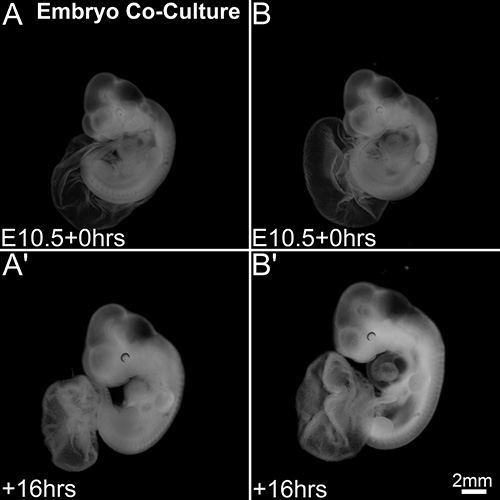
Figure 4. Developmental differences in mouse embryo coculture system. Embryos at the start of coculture (A, B). Embryonic development after 16 hr of coculture (A’, B’). One of the embryos in the coculture (A’) appears under-sized compared to the other embryo (B’) in the coculture. Scale bar in (B’) applies to all panels. Please click here to view a larger version of this figure.
| Total embryos cultured for 16-18 hr | Development after Culturing for 16-18 hr | Total embryos cultured for 38-40 hr | Development after Culturing for 38-40 hr | |||
| Good | Intermediate | Poor | Good | Poor | ||
| 112 | 65 | 31 | 16 | 12 | 4 | 8 |
Table 1. Development of embryos in mouse embryo culture system. Embryos cultured for either 16-18 hr or 38-40 hr. Of the total 112 embryos cultured for 16-18 hr, 65 (58%) embryos exhibited comparable development to embryos developing in utero, 31 (28%) showed intermediate and 16 (14%) showed poor development. Of the total 12 embryos cultured for 39-40 hr, only 4 (~35%) embryos showed good development while the remaining showed poor development.
