Recombinant human and porcine FVIII were successfully organized helically on negatively charged single bilayer LNT, resembling the activated platelet surface. The helical organization of the human and porcine FVIII-LNT was consistent through the collected digital micrographs (Figure 2). The control LNT and the human and porcine FVIII-LNT helical tubes were selected and segmented with the e2helixboxer.py GUI and initial data sets created with the e2workflow.py GUI, Single particle option (Table 1).
The helical order of the membrane-bound human and porcine FVIII-LNT was evaluated from the Fourier transform of the class averages with the e2display.py GUI (EMAN2) (Figure 3). The lipid bilayer in the best control LNT 2D class averages is well defined. The inner and outer leaflet and lower density of the membrane hydrophobic core are clearly visible (Figure 3A). The projected density of the membrane-bound human and porcine FVIII molecules oriented towards and perpendicular to the membrane surface is well defined and clearly shows the variations in the helical organization between the two proteins (Figures 3B and 3C). The more pronounced twist for the human FVIII-LNT helical tubes indicates that the protein-protein interactions between adjacent membrane-bound FVIII molecules are consistently different for the two FVIII forms (Figures 3B and 3C). Particles from class averages showing good helical organization (helical diffraction pattern) were merged in the e2display.py GUI to form an intermediate particle set (Table 1). The particles from the intermediate particle sets were again classified in 50 classes with the same constraints. The particles from class averages with the same diameter were merged in the final data sets (Table 1).
Initial 3D reconstructions for the human and porcine FVIII-LNT were carried out with 1,000 representative particles from the final human and porcine FVIII-LNT data sets. One hundred consecutive IHRSR iterations were run for each 3D reconstruction with a featureless cylinder (160 Å inner and 500 Å outer diameter), as initial volume. The axial rise ( Δz) calculated from the combined Fourier transform of the helical segments (particles set) is equal to 41 Å for human FVIII-LNT and 36 Å for porcine FVIII-LNT (Figures 4A and 4B). The initial azimuthal angle (ΔΦ) defined from the iterative search is estimated at 40.0° for the human FVIII-LNT and at 35.0° for the porcine FVIII-LNT. The final volumes are inspected for convergence of the helical parameters and correspondence between class averages and projections from the final reconstruction, also following the criteria described in5. The selected 3D reconstructions and corresponding helical parameters are imposed as initial volumes and initial helical parameters for a second IHRSR refinement of 100 cycles which converged to a four-start helical organization for the human FVIII-LNT with Δz = 41.1 Å and ΔΦ = 42.0° and a five-start helical organization for the porcine FVIII-LNT with Δz = 35.5 Å and ΔΦ = 34.8°. A final 100 IHRSR iterations imposing a 4-fold and a 5-fold helical symmetry for the human and porcine FVIII-LNT reconstructions respectively are carried out with initial volumes and corresponding helical parameters from the last asymmetric IHRSR refinements (Figures 4C and 4D). The final volumes show 8 human FVIII and 10 porcine FVIII membrane-bound molecules organized around the helical axis (Figure 5A). Each human FVIII molecule is translated 41.2 Å and rotated 42.0° from the previous one and each porcine FVIII molecule is translated 35.9 Å and rotated 35.2º from the previous one, corresponding to the helical parameters of the final 3D reconstructions (Figure 5B).
The reconstructed electron tomograms confirm the difference in the helical organization between the human and porcine FVIII-LNT obtained at the same experimental conditions. Comparison of the top views from the reconstructed tomograms and the 3D volumes from the helical reconstruction viewed in the direction perpendicular to the helical axis, further validates the correctness of the 3D reconstructions refined with the IHRSR helical parameters (Figure 6). The asymmetric 2D unit cell dimensions for the human FVIII-LNT 3D reconstruction are: a = 17.8 nm, b = 8.2, γ = 84° and for the porcine FVIII-LNT 3D reconstruction: a = 18.4, b = 7.2 and γ = 70° (Figure 6). The unit cell dimensions of human FVIII organized in membrane-bound 2D crystals are: a = 8.1, b = 7.0 and γ = 67º, which corresponds to the surface covered by one FVIII molecule viewed toward the membrane-surface20. Comparing the unit cell dimensions between FVIII organized in 2D and helical crystals indicates that both human and porcine FVIII molecules form dimers when helically organized on the LNT surface.
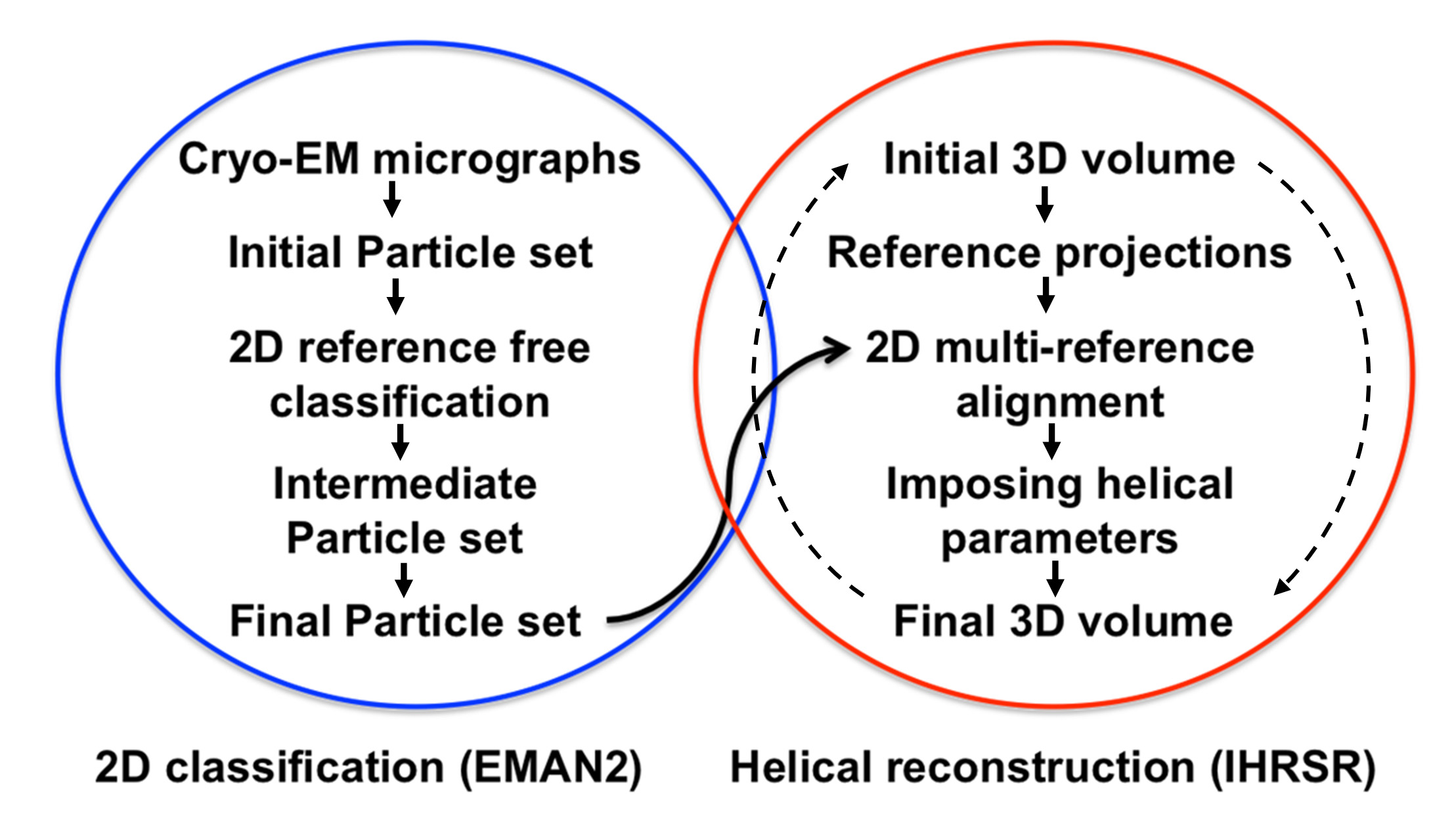
Figure 1. Structure analysis flow chart. The steps followed for the 2D classification analysis based on reference free alignment algorithms implemented in EMAN216 are circled in blue. The steps followed for the 3D analysis carried out with the iterative helical real space reconstruction algorithms (IHRSR) are circled in red. The iterative IHRSR cycles are denoted with dashed arrows.

Figure 2. Cryo-EM digital micrographs. (4,096 x 4,096 pixels, 2.9 Å/pix) of lipid nanotubes (LNT) with and without bound FVIII. A. Control LNT. B. Human FVIII-LNT. C. Porcine FVIII-LNT. The edge of the hole in the carbon film in which the FVIII-LNT are suspended in amorphous ice is indicated with a white star. The protein and lipid densities are in black. The magnified views (insets) of 512 x 512 cropped areas (white dashed square) illustrate the difference in the helical organization of the human and porcine FVIII, respectively. The scale bar is 100 nm. Please click here to view a larger version of this figure.

Figure 3. Representative 2D class averages (top row) and corresponding Fourier transforms (bottom row) from the intermediate particle sets (Table 1) classified in 50 classes. A. Control LNT B. Human FVIII-LNT C. Porcine FVIII-LNT. The class number and number of particles included in each class are indicated. The difference in helical order between the human and porcine FVIII is clearly seen on the images and confirmed by the diffraction patterns obtained from the Fourier transforms of these images. Please click here to view a larger version of this figure.
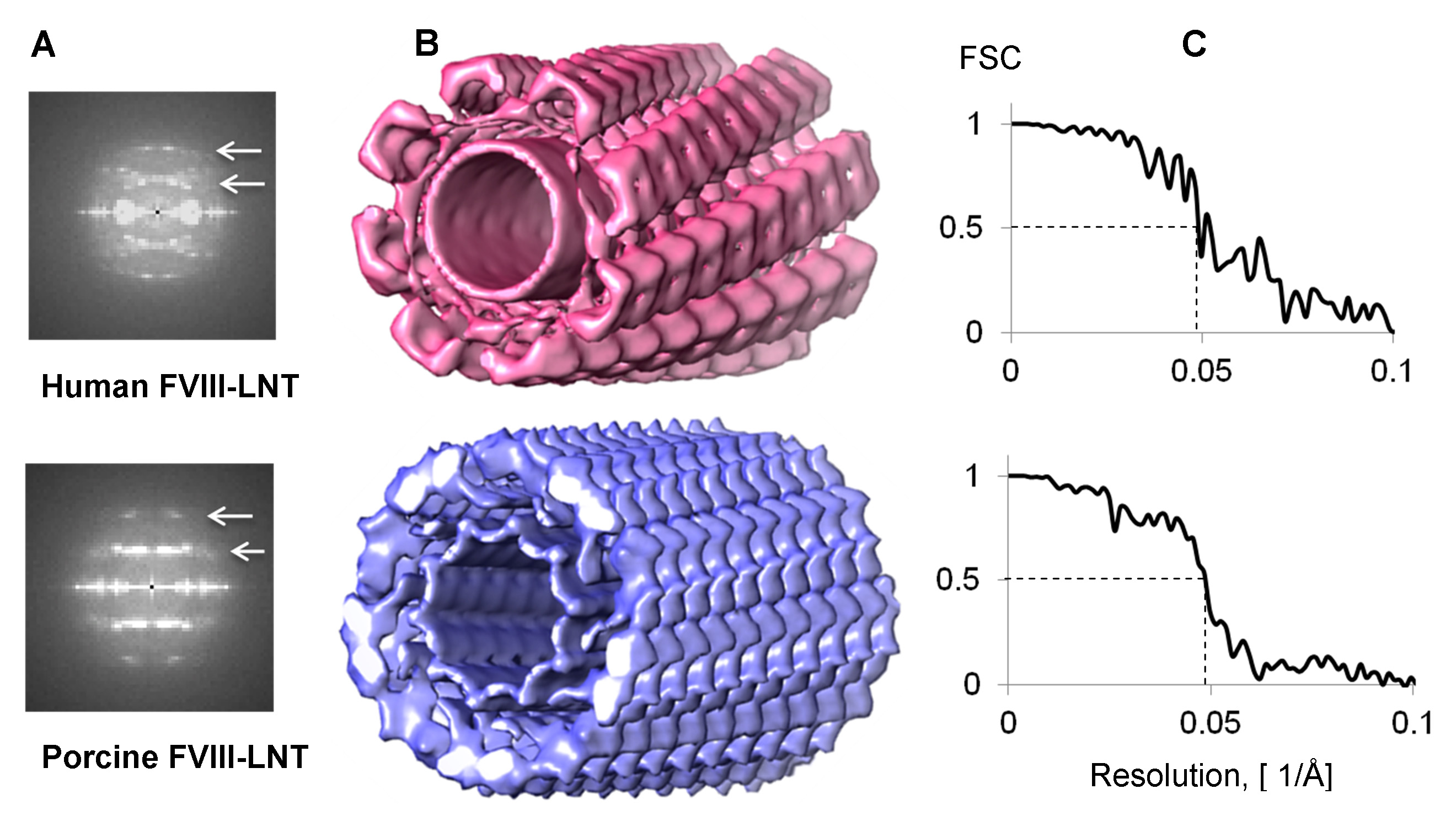
Figure 4. 3D helical reconstructions of human and porcine FVIII-LNT. A. Combined Fourier transform from 1,000 helical segments. The first and second layer line are centered at 1/82 Å-1 and 1/41 Å-1 for human FVIII-LNT, and at 1/72 Å-1 and 1/36 Å-1 for porcine FVIII-LNT (white arrows). B. Surface representation of human in pink (Δz = 41.1 Å, ΔΦ = 42.0º) and porcine in blue (Δz = 35.9 Å, ΔΦ = 35.2º) FVIII-LNT 3D helical reconstructions. Both volumes are presented at 0.005 contour level (minimum density is 0 and maximum density 0.02, as calculated in UCSF Chimera, Volume viewer option21). The length of the FVIII-LNT tube is 256 pixels at 2.9 Å/pix. C. Fourier Shell Correlation (FSC) plots for human and porcine FVIII-LNT showing a resolution of 20.5 Å at FSC = 0.5.
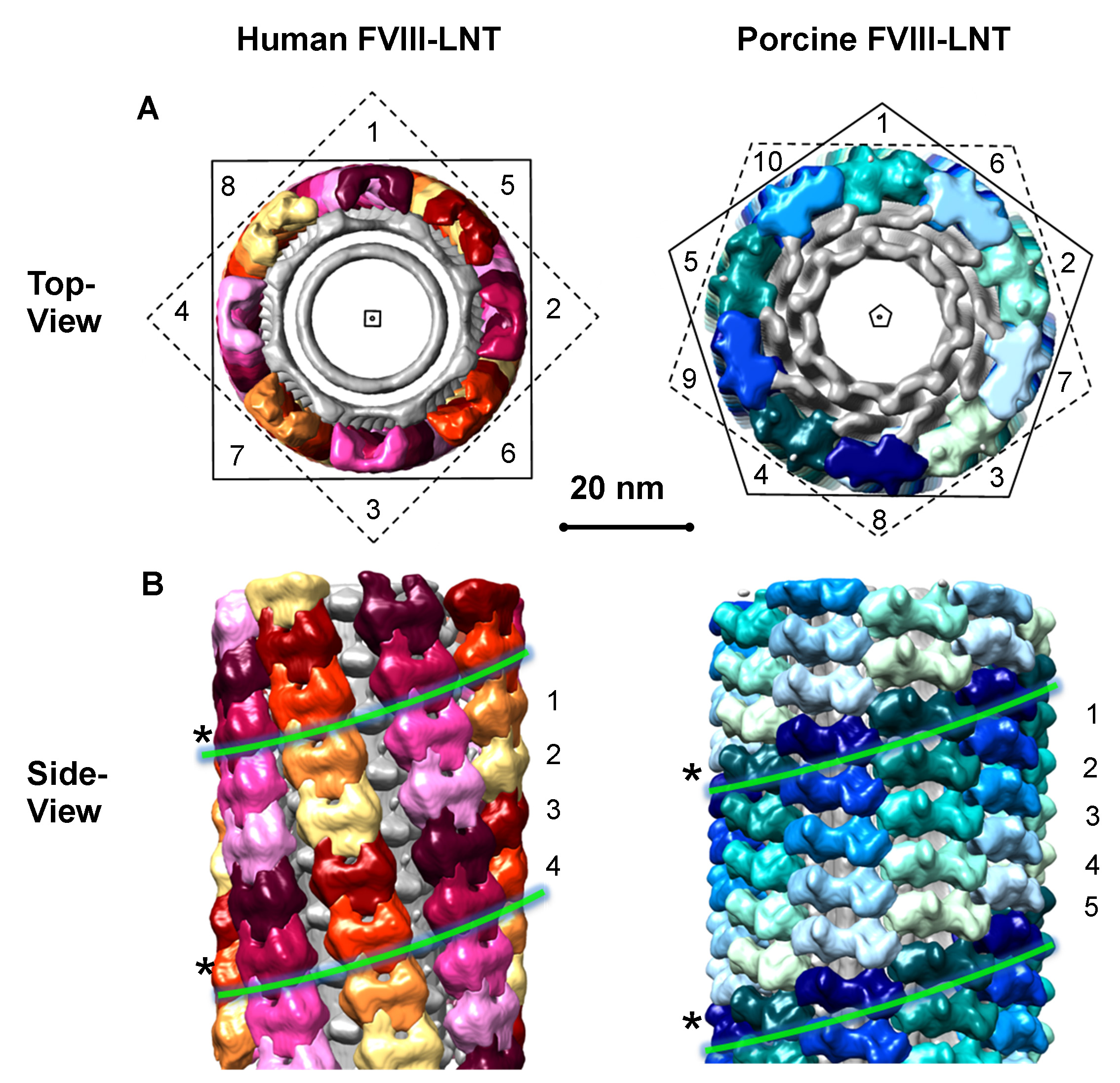
Figure 5. Helical organization of human and porcine FVIII-LNT. Segmented surface representation of human and porcine FVIII-LNT helical reconstructions, shown in Figure 4B. The volumes are segmented after imposing 4-fold symmetry to the human FVIII-LNT and 5-fold symmetry to the porcine FVIII-LNT. The asymmetric units are color coded yellow-red for the human FVIII-LNT and blue-green for the porcine FVIII-LNT. A. Views along the helical axis indicated with a square for the human and as a pentagon for the porcine FVIII-LNT. The human FVIII-LNT structure shows 8 molecules organized around the outer LNT membrane and the porcine FVIII structure shows 10 molecules organized around the outer LNT membrane, indicated with numbers. B. Views perpendicular to the helical axis. The human FVIII-LNT is a 4-start helical structure and the porcine FVIII-LNT is a 5-start helical structure. The individual one start helices are indicated with numbers and color-coded. We have emphasized one of the helices from each structure with a (*) and green lines. The scale bar is 20 nm.
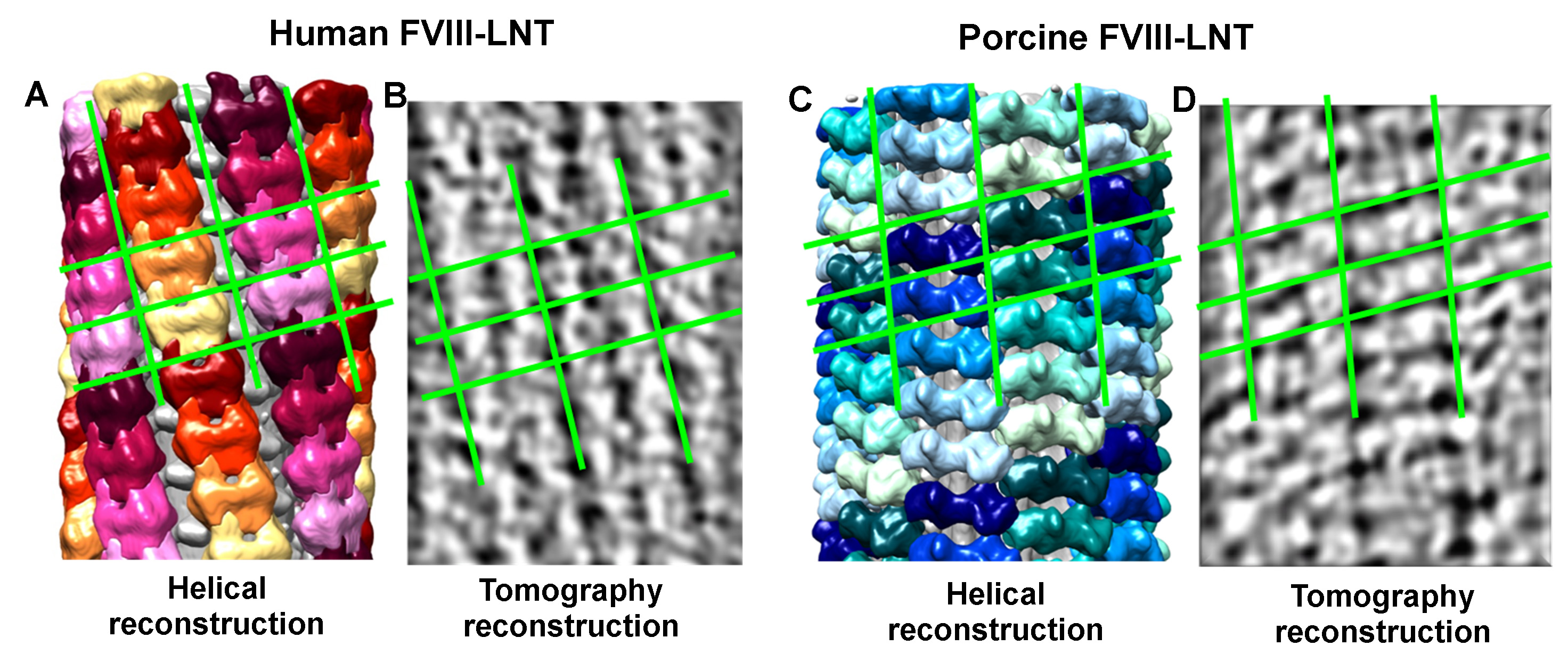
Figure 6. Comparison between the helical and tomography 3D reconstructions. The human FVIII-LNT (A) and porcine FVIII-LNT (C) helical 3D reconstructions are shown perpendicular to the helical axis. Each unit cell and individual helices are color-coded as in Figure 5. B. and D. are density representations of the 3D tomography reconstructions, viewed perpendicular to the helical axis. The 2D lattice reflecting the helical arrangement of the FVIII molecules is shown with green lines.
| SAMPLES | cLNT | hFVIII-LNT | pFVIII-LNT |
| Initial Micrographs | 61 | 474 | 542 |
| Initial Particle sets | 29113 | 60395 | 64665 |
| Defocus (nm) | -4,051 ± 502 | -3,643 ± 737 | -3,443 ± 1,086 |
| Intermediate Particle sets | 25,907 | 27,305 | 22,773 |
| Final Particle sets | 25,907 | 10,455 | 10,430 |
Table 1. 2D analysis statistics following the algorithm presented in the flowchart on Figure 3.
