A primary screen tested 328 ORs against 26 odors at a concentration of 100 µM. This odor concentration has been demonstrated to effectively activate a large proportion of ORs with known ligands10. First, normalized luciferase activity was calculated by dividing the firefly luciferase reading by the Renilla luciferase reading. Next, baselined values were calculated by subtracting the normalized luciferase readings for the no odor control from the normalized luciferase readings for each well (Figure 1). Dose response curves were performed on 48 odorant/OR pairs randomly distributed across the range of baselined values, as indicated by colored bars in Figure 1. ORs were treated with 7 concentrations of odorant spanning 1 nM to 1 mM, and the resulting responses were fit to a sigmoidal curve using nonlinear regression. An odorant/OR was considered an agonist if it met three criteria: (1) the standard error of the logEC50 was less than 1 log unit; (2) the 95% confidence intervals for the top and bottom parameters of the curve did not overlap; and (3) the extra sums-of-squares test confirmed that the odorant activated the OR-containing cells significantly more than the control cells, which were transfected with an empty vector. Dose response results are summarized in Table 3.
These data were then used to determine how well assay measurements in a primary screen predict results from the dose response curve. Blue bars in Figure 1 correspond to pairs that were classified as agonists in a full dose response experiment, while red bars did not meet our three criteria outlined above. Values from the primary screen predicted results from the full dose response experiment (area under the receiver operating characteristic curve (AUC) = 0.68, p < 0.01, Mann-Whitney U test), indicating that our primary screen is a useful method to enrich for odorant/OR pairs that will be classified as agonists in a full dose response experiment (Figure 2).

Figure 1. Frequency of baselined luciferase values for a screen with a panel of olfactory receptors and odorants. Histogram of the frequency (Count) of baselined luciferase values calculated for each odorant/OR pair in the primary screen. As odorant/receptor activation pairs are sparse, the majority of the values are centered at zero and the large central distribution estimates the noise distribution for this assay. Colored bars indicate odorant/receptor pairs chosen for dose response analysis; blue bars are pairs that were classified as agonists based on the full dose response, and red bars are pairs that were not classified as agonists. Click here to view larger image.
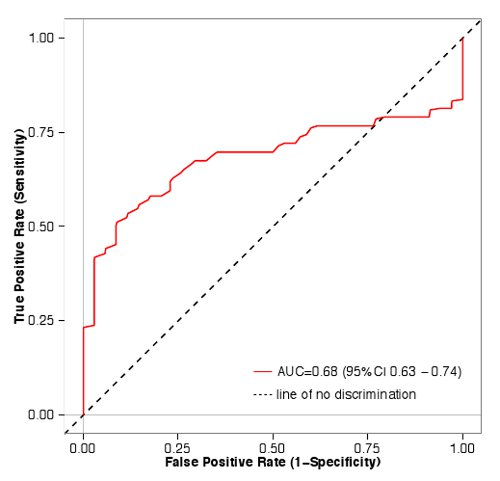
Figure 2. ROC curve for the odorant/receptor screen. 48 odorant/receptor pairs were classified as being agonists or as not being agonists. True positive rate (sensitivity) was then plotted against the false positive rate (1-specificity) using the R statistical package40. The area under the curve (AUC) is 0.68, indicating that odorant/receptor pairs with higher luciferase screen values are more likely to pass dose response than those with lower values. Click here to view larger image.
| Baselined Value | Dose Response |
| 0.051793067 | fail |
| 0.006376956 | fail |
| 0.331936398 | pass |
| 0.591006519 | pass |
| 0.049093369 | pass |
| 0.396788976 | pass |
| -0.013655743 | pass |
| 0.011080217 | pass |
| 0.004203349 | fail |
| 0.003975049 | fail |
| -0.077935718 | pass |
| -0.084488317 | pass |
| 0.030236078 | fail |
| -0.042963576 | fail |
| 0.031466406 | fail |
| 0.025897747 | fail |
| -0.030434651 | fail |
| -0.004122795 | fail |
| -0.010075533 | fail |
| 0.028883452 | fail |
| 0.019402373 | fail |
| 0.047508749 | fail |
| 0.00255344 | fail |
| 0.017221449 | fail |
| 0.340216655 | pass |
| -0.026912181 | fail |
| 0.037140428 | fail |
| 0.467763017 | pass |
| 0.097665337 | fail |
| 0.080657267 | pass |
| 0.172819211 | pass |
| 0.05568393 | pass |
| -0.106721064 | pass |
| 0.136614849 | pass |
| 0.457839849 | fail |
| 0.211751741 | fail |
| 0.1581464 | pass |
| -0.62099155 | pass |
| -0.066949491 | pass |
| -0.78712035 | pass |
| 0.752503007 | pass |
| 1.433407558 | pass |
| 0.475431098 | pass |
| 1.457936815 | pass |
| 0.048652537 | fail |
| 0.027196782 | fail |
| 0.129599842 | fail |
| -0.069781272 | fail |
| 0.016450039 | fail |
| -0.025639207 | fail |
| 0.158152141 | fail |
| -0.032570055 | fail |
| 0.140139926 | fail |
| -0.052030276 | fail |
| 0.657140133 | pass |
| 1.040410297 | pass |
| 0.164647156 | pass |
| 0.399588712 | pass |
| 0.188094387 | pass |
| 0.039371424 | pass |
| 0.016784352 | pass |
| 0.229959571 | pass |
| 0.238381997 | fail |
| 0.074118909 | fail |
| 0.423901128 | pass |
| 0.152621022 | pass |
| -0.109048046 | pass |
| 0.075301806 | pass |
| 0.395233972 | pass |
| 0.261892958 | pass |
| 0.156693306 | fail |
| 2.163418147 | pass |
| 3.649862104 | pass |
| 0.025716169 | pass |
| -0.033258008 | pass |
| -0.026984127 | fail |
| -0.338441868 | pass |
| 0.37398618 | pass |
Table 3. Olfactory receptor/odor pairs tested in dose response. Baselined luciferase values and dose response results (pass or fail) for 48 OR/odor pairs chosen from the screen. For 30 pairs tested in the screen twice, both baselined luciferase values are included.
