C26 tumor growth kinetics show a lag phase for the first 7 – 8 d after injection, followed by exponential cell growth (4 – 5 d). The tumor mass eventually reaches ~10% of the body weight (about 2 g; Figure 1A-B). During the first phase, the tumor can be located by palpation only and appears as a small protrusion of the skin. In the second phase, the tumor is observed as a mass under the skin. Rarely, the tumor becomes ulcerated, resulting in an open wound; in this case, the mouse is excluded from the experimental group and is humanely euthanized.
Body weight is unchanged in the first phase, but it is significantly reduced in the second phase, when it reaches 10 – 15% of the initial body weight (30% in the case of tumor-free weight; Figure 1A). Tumor-bearing mice appear wasted and showed disheveled fur at the end of the experimental period, with a body condition (BC) score equal to 118 (Figure 1C). BC1 represents a severely emaciated mouse, where skeletal structures are evident and the vertebrae are distinctly segmented. Body weight loss is mainly accounted for by both skeletal muscle and fat tissue wasting (Figure 1B). Body weight loss is consistent with a reduction of about 20 – 30% in skeletal muscle weight, in particular in the gastrocnemius, tibialis anterior, and quadriceps (Figure 2). The cardiac muscle is also significantly reduced in weight, although to a lesser extent when compared to the other skeletal muscles (Figure 2). Interestingly, hepatomegaly (+16%, p < 0.01) and splenomegaly (+110%, p < 0.01) are generally detected in tumor hosts, while fat mass, similar to skeletal muscle, is severely depleted (-70%, p < 0.001; Figure 3).
Skeletal muscle weight loss is also consistent with and proportional to the reduction in muscle fiber size, as observed after morphometric evaluation of muscle fiber CSA by means of an IF method (Figure 4A-B). In particular, the frequency distribution analysis showed a shift towards smaller-size fibers in C26-bearing mice, thus suggesting that the whole muscle undergoes atrophy in the presence of a C26 tumor (Figure 4C). Similar results can be observed by taking advantage of a traditional H&E-based methodology for the quantification of fiber size, although the magnitude of the change in muscle CSA associated with cancer growth is slightly different (38% versus control, p < 0.01 for the IF-based method; -18% versus control, p < 0.01 for the H&E-based method; Figure 5).
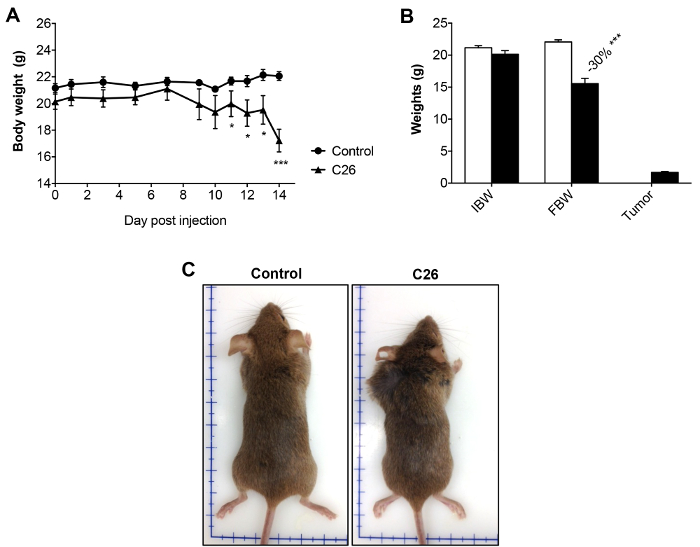
Figure 1: Body Weights in Control and C26-bearing Mice. A) Body weight curve in control and C26 hosts over the entire experimental period (14 days after tumor injection). B) Initial body weight (IBW), final tumor-free body weight (FBW), and tumor weight in control and C26-bearing animals. C) Representative pictures of controls and C26-bearers at the time of sacrifice (day 14 after tumor inoculation). Data are expressed as the mean SD. n = 6. Significance of the differences: *p < 0.05, **p < 0.01, ***p < 0.001 versus the control group using Student's t-test. Please click here to view a larger version of this figure.
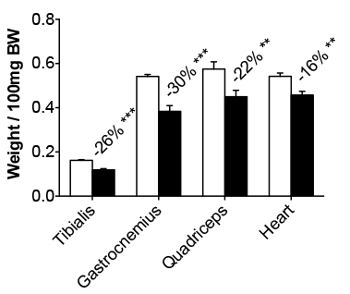
Figure 2: Muscle Weights in Control and C26-bearing Mice. The tibialis, gastrocnemius, quadriceps, and heart weights in control and C26 tumor-bearers are reported as the weight/100 mg IBW. The data are expressed as the mean SD. n = 6. Significance of the differences: **p < 0.01 and ***p < 0.001 versus the control group using Student's t-test.
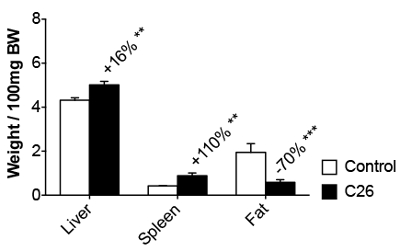
Figure 3: Organ Weights in Control and C26-bearing Mice. The liver, spleen, and epididymal fat pad weights in control and C26 tumor-bearers are reported as the weight/100 mg IBW. The data are expressed as the mean SD. n = 6. Significance of the differences: **p < 0.01 versus the control group.
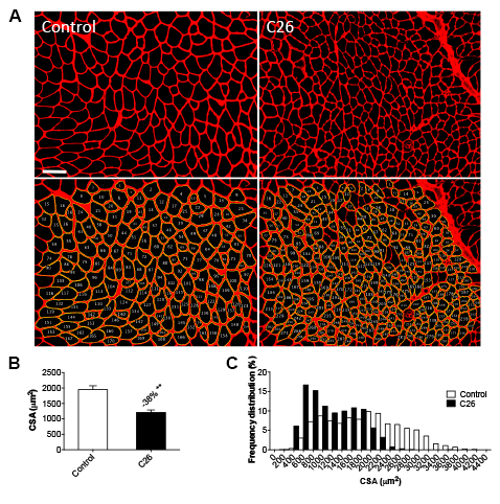
Figure 4: Morphometry for Myofiber Size by Immunofluorescence. A) Fluorescent immunohistochemical reaction of tibialis sections using an anti-dystrophin antibody (Vector Laboratories) from control and C26 hosts. Magnification: 10X. Size bar: 100 µm. B) Quantification of CSA in the tibialis muscle measured with the ImageJ macro. C) Frequency distribution of CSA in the tibialis muscle of control and C26-bearing mice. The data are expressed as the mean ± SD. n = 6 for each group. Significance of the differences: **p < 0.01 versus the control group using Student's t-test.
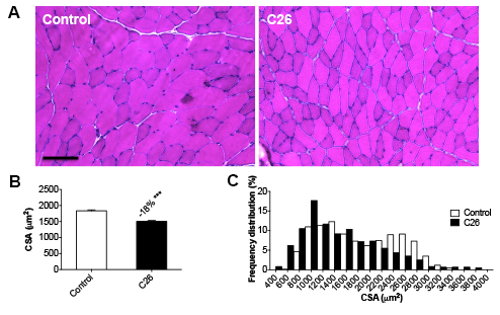
Figure 5: Morphometry for Myofiber Size by H&E. A) Hematoxylin & eosin staining of tibialis sections from control and C26 hosts. Magnification: 20X. Size bar: 100 µm. B) Quantification and frequency distribution of CSA in the tibialis muscle. The data are expressed as the mean SD. n = 6. Significance of the differences: ***p < 0.001 versus the control group using Student's t-test.
| Autori | Mouse strain | Mouse gender | Cell number | Origin | Implanted as solid tumor? | Site of injection | Days |
| Lazarus et al., 1999 | CD2F1 | M | 5 x 105 | NCI | NO | Dorsal region | 17 |
| Al-Majid and McCarthy, 2001 | CD2F1 | F | 2.5 x 106 | OSUMC | NO | Dorsal region | 17 |
| Samuels et al., 2001 | Balb/c | M | 0.5 g/ml of tumor suspension | ns | YES | Dorsal region | Up to 11 |
| Acharyya et al., 2004 | CD2F1 | M | 1 x 107 | OSUMC | NO | Right flank | Up to 24 |
| Aulino et al., 2010 | Balb/c | ns | 0.5 mm3 | NCI | YES | Dorsal region | 21 |
| Benny-Klimek et al., 2010 | CD2F1 | F | 1 x 106 | OSUMC | NO | Dorsal region | 14 |
| Bonetto et al., 2011; 2012 | CD2F1 | F | 1 x 106 | OSUMC | NO | Dorsal region | 14 |
| Cosper and Leinwand, 2010 | CD2F1 | M+F | 5 x 105 | ns | NO | Right flank | Up to 27 |
| Murphy et al., 2012 | CD2F1 | ns | 5 x 105 | NCI/OSUMC | NO | Dorsal region | 14 |
| Cornwell et al., 2014 | CD2F1 | M | 5 x 105 | NCI | NO | Right&Left flank | Up to 26 |
| Assi et al., 2016 | Balb/c | ns | 1 x 106 | Cell Line Service | NO | Dorsal region | Up to 22 |
Table 1: Comparison of the Different Protocols used for C26 Implantation. Different protocols for C26 implantation are provided, with particular reference to the mouse strain used, the cell number and type of tumor inoculated, the cell origin, the site of injection, and the length of the experimental period. OSUMC: Ohio State University Medical Center. NCI: National Cancer Institute. Commercial cell line service NS: not specified.
