Targeting of ChR2 to GABAergic cells in the MS as described in section 1 is illustrated in Figure 2A. Optogenetic stimulation of axons of MS GABAergic cells in the dorsal hippocampus via an optic fiber which is implanted above the CA1 area entrains theta oscillations at the frequency of the stimulus in the ipsilateral (Figure 2B) as well as contralateral hemisphere (Figure 2C). Theta oscillations could be more or less efficiently entrained by the optogenetic stimulation (Figure 3A), the efficacy of which was computed for each recording epoch as a relative theta LFP power around the stimulation frequency, i.e., entrainment fidelity (Figure 3B). Entrainment fidelity above 0.3, i.e., higher than in the spontaneous light-off recordings, was observed in approximately 80% of the recording epochs. Optostimulation at non-theta frequencies was less effective (Figure 3C).
Explicit i.e., parametric manipulation of theta oscillations frequency is accompanied by emergent changes of theta regularity: the temporal regularity of amplitude and frequency of the theta oscillations was increased during epochs with high entrainment fidelity. The stability of oscillations can also be regulated parametrically by applying trains of light pulses, periods of which follow Gaussian distributions with different dispersions (Figure 4).
Optogenetic control over the oscillations frequency eliminated the correlation between theta frequency and running speed, in agreement with the frequency control via the MS by ascending afferents during movement (Figure 5A). Optostimulation also induced theta oscillations during immobility (Figure 5B). The preferential firing phases recorded in the CA1 area in putative pyramidal cells and interneurons were unchanged relative to the optogenetically entrained theta oscillation when compared to spontaneous theta (Figure 6).
To study the contribution of the hippocampus to the lateral septum pathway in theta-mediated regulation of locomotion, we optogenetically inhibited this pathway. Halorhodopsin (eNpHR3.0) was bilaterally expressed in hippocampal pyramidal cells (Figure 7A), whereas ChR2 was expressed in MS GABAergic cells as above and theta oscillations were optogenetically entrained (Figure 7B). The theta entrainment reduced variability of running speed but not when the hippocampus to the LS pathway was inhibited (Figure 7C).
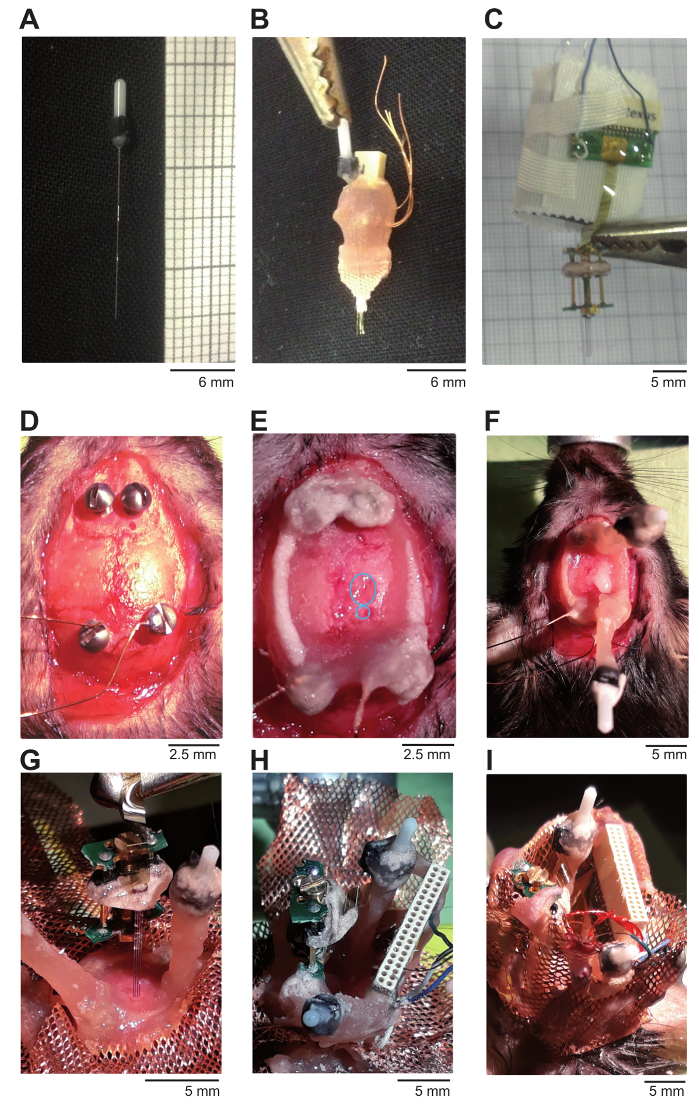
Figure 1: Illustration of optic fibers, electrodes and surgery. (A) Illustration of an optic fiber. (B) Illustration of a wire array glued to an optic fiber for the recording of hippocampal LFP during entrainment of hippocampal theta oscillations. (C) For recording of the hippocampal cellular activity, a silicone probe is mounted on a microdrive. (D) Miniature screws are positioned on the skull. Copper wires are presoldered to the ground and reference screw before positioning them above the cerebellum. (E) Cement is applied to cover and connect the screws. The upper blue circle indicates where the craniotomy was performed for the implantation of the silicone probe. Lower blue circle indicates where the craniotomy was performed for the implantation of the optic fiber in the hippocampus. (F) One optic fiber is implanted in a caudal-rostral angle to target the hippocampal CA1 region. A second fiber can be implanted in the medial septum if stimulation of cell somata is desired (optional). (G) The silicone probe is lowered to just above the hippocampal CA1 area. (H) The borders of the microdrive and connector are cemented to the implant and ground, and the reference wires are soldered. (I) Copper mesh is constructed to surround the implant and serve as a Faraday cage. Please click here to view a larger version of this figure.
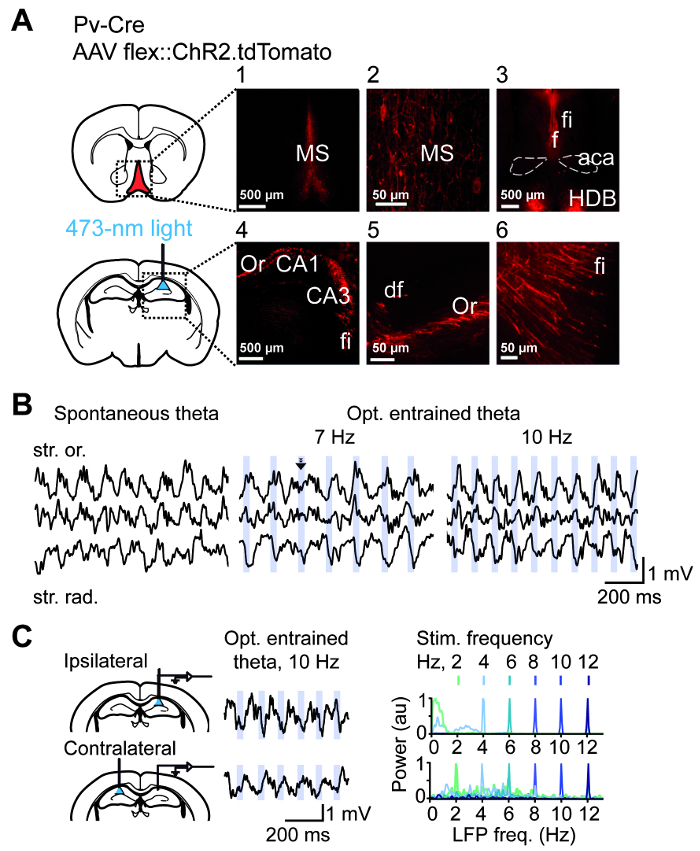
Figure 2: Preparation for optogenetic hippocampal theta entrainment. (A) ChR2 was expressed in PV+ medial septal cells in PV-Cre mice (upper scheme). Bright fluorescence in MS (1, 2) confirms successful construct expression in somata. MS fibers project via fornix (f) and fimbria (fi) to the hippocampus (3-6); aca: anterior commissure; anterior part. HDB: nucleus of the horizontal limb of the diagonal band; Or: stratum oriens. The optic fiber for optogenetic stimulation with blue light is implanted above the pyramidal layer of hippocampal area CA1 (lower scheme). Scale bars: 500 µm (images 1, 3, 4) and 50 µm (images 2, 5, 6). (B) Hippocampal LFP during spontaneous theta oscillations (left) and 7 Hz (middle) or 10 Hz (right) optogenetic entrainment. Blue stripes indicate the time windows of light application. Note the phase reset by the light pulse indicated by an arrow. Note gamma envelopes during spontaneous and entrained theta, an indicator of physiological theta rhythm. Phase reversal between stratum oriens (str. or.) and stratum radiatum (str. rad.) is also maintained during entrainment. (C) Entrainment is reliable during ipsilateral (upper plots), as well as contralateral (lower plots) optogenetic stimulation. Schemes illustrate positions of fibers in respect to electrodes positions. Example LFP traces during theta and application of light pulses are shown in the middle. On the right, power spectra of hippocampal LFP during ipsi- and contralateral stimulation color-coded according to stimulation frequency. This figure has been modified from ref. 11. Please click here to view a larger version of this figure.
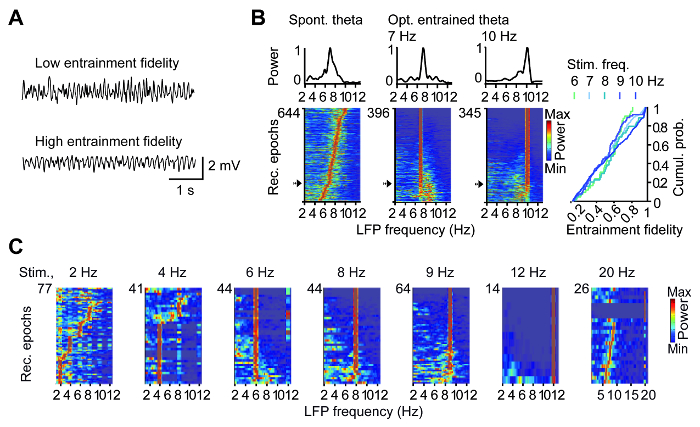
Figure 3: Fidelity of optogenetic hippocampal theta entrainment. (A) Example hippocampal LFP traces during low and high entrainment fidelity. (B) Power spectral densities of 10 s epochs during spontaneous theta, with rows ordered according to leading theta frequency (left), and during 7 Hz (middle) and 10 Hz (right) optogenetic stimulation, with rows ordered according to entrainment fidelity. Respective example power spectra (indicated by an arrow) are plotted above. Note reliable entrainment fidelity across epochs. On the right, the cumulative probability of entrainment fidelity for theta frequencies is shown. (C) Entrainment requires theta rhythmic stimulation. Hippocampal network activity can be successfully entrained using frequencies between 6-12 Hz. At lower frequencies (e.g., 2 or 4 Hz) or higher frequencies (e.g., 20 Hz) entrainment is not reliable. This figure has been modified from ref. 11. Please click here to view a larger version of this figure.
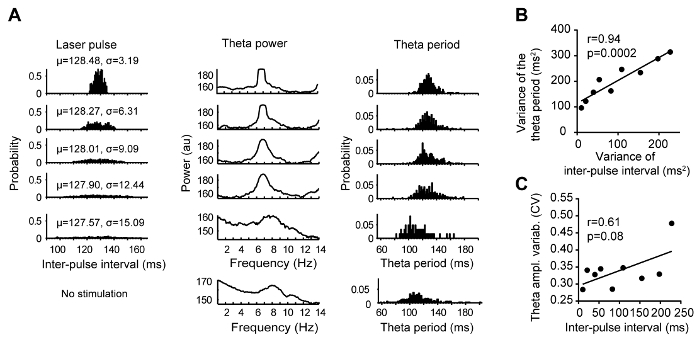
Figure 4: Parametric manipulation of theta oscillations regularity. (A) Stimulation was applied at varying frequencies within the theta range with a mean frequency of 7.8 Hz following a Gaussian distribution. The standard deviation of the inter-pulse intervals was increased across protocols from σ = 3.19 to σ = 15.09. In total, 11 protocols were generated and applied, each with a total duration of the stimulation epoch of 1 min. Of those, the probability distribution of 5 protocols are shown on the left of the figure. The power spectral densities within a range of 1-14 Hz of the hippocampal LFP during application of the respective protocols are plotted in the middle of the figure. The probabilities of the theta periods during application of the respective protocols are illustrated on the right. (B) The variance of the applied inter-pulse intervals determined the variance of the concurrent theta period (Pearson's r = 0.94, p = 0.0002). (C) The relationship between the theta amplitude variability and the inter-pulse interval (Pearson's r = 0.61, p = 0.08). This figure has been modified from ref. 70. Please click here to view a larger version of this figure.
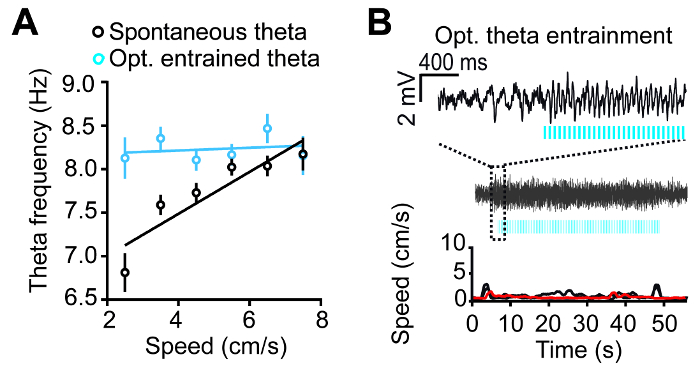
Figure 5: Optogenetic theta rhythmic entrainment determines hippocampal LFP during behavior. (A) Optogenetic stimulation frequency determined the theta frequency during locomotion. Hence, speed-related afferents do not impact hippocampal theta frequency, and as a consequence, speed is not correlated with theta frequency (blue) as it is during spontaneous theta (black). Data are presented as mean ± s.e.m. (B) During quiet wakefulness, the hippocampal theta can be elicited in the absence of movement. Hippocampal LFP traces before and during successful entrainment are shown above, and example speed traces recorded during entrainment are shown below (the red trace corresponds to the hippocampal LFP trace depicted above). Blue stripes mark the time windows of light stimulation pulses. This figure has been modified from ref. 11. Please click here to view a larger version of this figure.
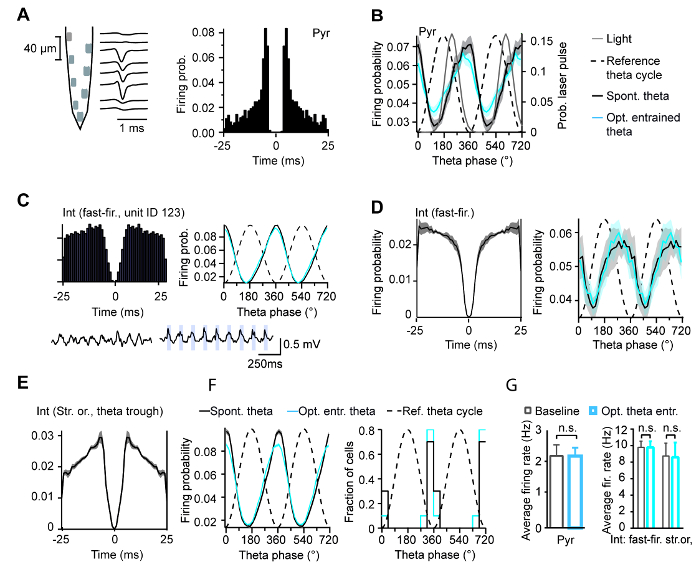
Figure 6: Hippocampal cellular activity during theta entrainment. (A) Cellular activity was recorded using silicone probes (scheme). Single interneurons and pyramidal cells were isolated and identified according to their respective waveform. Shown here is the average waveform (middle) and auto-correlogram of an example isolated pyramidal cell. (B) Preferred discharge phase of pyramidal cells (Pyr) was not different during spontaneous (in black, n = 29 neurons) and optogenetically entrained (in blue, n = 30) theta (p = 0.79). (C) Shown here is the auto-correlogram (left) and preferential firing phase of a fast-firing interneuron during spontaneous and optogenetically entrained theta. Below the corresponding hippocampal LFP rhythm during spontaneous (left) and entrained (right) theta. (D) Preferred discharge phase of fast-firing interneurons was not different during spontaneous (in black) and optogenetically entrained (in blue, n = 28 neurons) theta (p = 0.97). Average auto-correlogram is shown on the left. (E) Average auto-correlogram str. oriens cells. (F) Preferred discharge phase of str. oriens interneurons was not different during spontaneous (black) and optogenetically entrained (blue, n = 10 neurons) theta (p = 0.56). Histograms of preferred discharge phases are shown on the right. (G) Average firing rates were not affected by theta entrainment in pyramidal cells (p = 0.98), fast-firing interneurons (p = 0.96) or str. oriens interneurons (p = 0.85). This figure has been modified from ref. 11. Please click here to view a larger version of this figure.
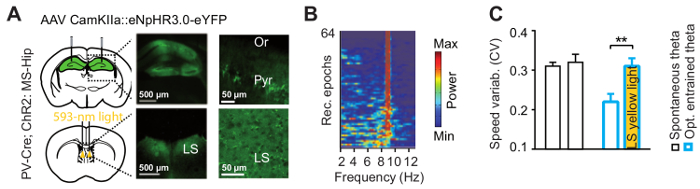
Figure 7: Combination of hippocampal theta entrainment and optogenetic inhibition of the hippocampal subcortical output through the LS. (A) eNpHR3.0 (halorhodopsin) was expressed in hippocampal pyramidal cells (upper scheme). Successful expression of the construct was confirmed by bright fluorescence in somata in the hippocampus (upper images) and axons in the LS (lower images). Optic fibers were implanted bilaterally above the LS (lower scheme). Scale bars: 500 µm (images on the left), 50 µm (images on the right). (B) Hippocampal theta is successfully entrained during inhibition of the hippocampus to LS pathway. Here shown are power spectral densities for 9 Hz blue light stimulation during output inhibition. (C) Inhibition of the major hippocampal subcortical output pathway prevents effects of hippocampal theta entrainment on speed. Here shown is decrease in speed variability upon optogenetic entrainment (white bar with blue borders), with is absent upon simultaneous inhibition of the hippocampus to LS pathway (yellow bar with blue borders). Respective average baseline speed is shown on the left. This figure has been modified from ref. 11. Please click here to view a larger version of this figure.
