The microtray-based single-worm culture environment described here can be used to monitor a variety of phenotypes, including lifespan and health span, activity and movement, body shape and crawling geometry, and the expression of transgenically expressed fluorescent biomarkers in individual animals over time. The microtray culture system is compatible with lifespan analysis through either manual scoring or image collection and downstream imaging analysis. As with standard culture on Petri plates21, worms can be manually scored as dead or alive based on movement following a stimulus (e.g., tapping the plate or exposure to blue light). For image-based lifespan analysis, a comparison of frame-to-frame differences between images taken after the stimulus has been applied is used to determine when each worm stops moving. This is compatible with automated image collection and analysis systems and provides additional data in the form of the activity level of individual animals at each time point. This information can then be used not just to estimate lifespan (cessation of movement), but also health span (a threshold of reduced activity; multiple definitions have been proposed). Additional parameters (body size, shape, and posture) can also be measured from these images.
The key features of this microtray culture system are (1) the capacity to track individual animals over time, (2) the compatibility between the culture system and aversive interventions like dietary restriction or chemical stress agents, and (3) the capability to measure fluorescence directly in the culture environment. To demonstrate these features, we exposed worms to 250 µM copper sulfate (CuSO4) to induce heavy metal stress or 10 mM dithiothreitol (DTT) to induce protein misfolding stress and measured daily activity, lifespan, and health span. Both CuSO4and DTT significantly reduced worm lifespan (Figure 2A). Monitoring daily activity allowed us to estimate the individual health span of each worm, here defined as the age at which a worm can no longer move a full body length in response to the stimulus. CuSO4 and DTT reduced health span relative to untreated controls (Figure 2B). In contrast, CuSO4 decreased the proportion of life spent in good health, while DTT did not (Figure 2C). CuSO4 also reduced average activity across individuals within the population to a greater extent than DTT at all ages (Figure 2D), as well as cumulative activity for each animal across the lifespan (calculated as the area under the activity curve [AUC]; Figure 2E). An important advantage of this microtray setup is its ability to correlate phenotypes measured at different time points across individual animals within a population. In this case, we find that cumulative activity for individual animals up to day 10 correlates with lifespan for DTT-challenged animals but not for untreated controls or CuSO4-challenged animals (Figure 2F).
To demonstrate the capability to monitor fluorescent biomarkers across the lifespan, we used the microtray culture system to monitor the expression of mScarlet transgene fused to the C-terminus of the kynu-1 gene, encoding the enzyme kynureninase in the kynurenine metabolic pathway, at its endogenous locus. We previously found that expression of human kynureninase, encoded by Kynu, increases with age in blood26, and that knockdown of kynu-1 di C. elegans is sufficient to extend lifespan27. KYNU-1 displays age-dependent expression; it is nearly undetectable at the L4 larval stage and peaks at day 9 of adulthood. A drastic increase in expression is observed between days 3 and 8 of adulthood, and there is a progressive decline with age thereafter (Figure 3A, Supplementary Figure 1). To assess the relationship between KYNU-1 and C. elegans aging, we quantified the cumulative abundance of KYNU-1 up to day 14 of adulthood (an age at which most animals were still alive) and found a significant correlation with lifespan (Figure 3B). Since KYNU1 expression is dynamic over the course of a lifespan, we compared KYNU-1 expression at different ages to overall survival across individual animals. At no single time point was KYNU-1 expression significantly correlated with survival (Figure 3C-F), demonstrating that lifelong expression is a better indication of lifespan impact than single time point assessments.
Combined, these experiments provide representative data that highlight the capabilities of the microtray-based single-worm culture system to enable dynamic changes in worm activity, health, lifespan, and fluorescent biomarkers to be captured and compared across ages in individual animals. In particular, the ability to monitor individuals at multiple time points allows dynamic changes in both physiological and molecular phenotypes to be tracked in vivo, enabling the detection of patterns that would not be apparent using cross-sectional measurements across populations.
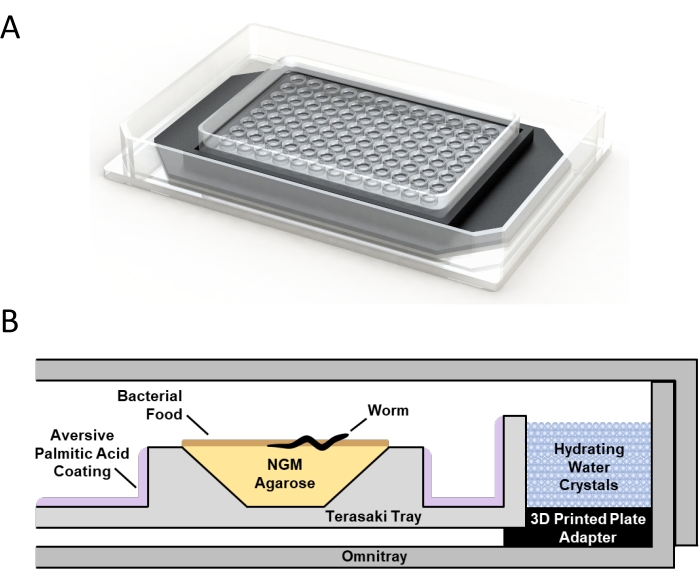
Figure 1: Microtray single-worm culture environment. (A) 3D rendering of a microtray mounted within a single-well plate using a custom 3D-printed adapter. (B) Schematic of the single-worm culture system showing the relative position of a microtray well with low-melt agarose nematode growth media (NGM), bacterial food, isolated C. elegans, aversive palmitic acid coating, the 3D printed adapter, water crystals, and the single-well plate. Please click here to view a larger version of this figure.
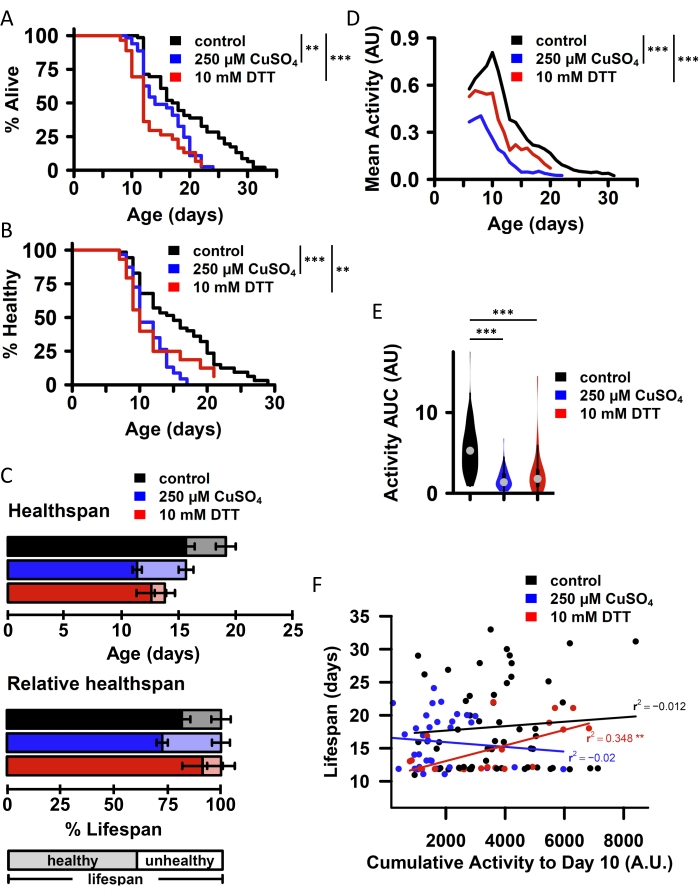
Figure 2: Correlation of phenotypes in individual animals across populations using Microtray single-worm environments. All panels provide data from an experiment comparing three group of wild-type (N2) C. elegans exposed to no additives (control; N = 55), 250 µM copper sulfate (CuS04; N = 38), or 10 mM dithiothreitol (DTT; N = 34) starting at day 2 of adulthood. Lifespan (A) and health span (B) are both moderately reduced by chronic exposure to either CuSO4 or DTT. Health span is defined as the last day an animal can move at least one full body length on the plate. Pairwise significance is calculated using the log-rank test (R survdiff function). (C) Health span and lifespan averages within each population (top: absolute values; bottom: normalized to each group's mean lifespan). (D) Average worm activity (3 day rolling mean of daily activity) within each group across the lifespan and (E) the population average of the individual animal area under the lifetime activity curve (AUC) are both substantially impaired by CuSO4 or DTT. Significance determined by the Mann-Whitney U test to compare AUC for individual animals between groups. (F) Cumulative activity for individual animals to day 10 (bottom) correlates with a lifespan for animals exposed to DTT, but not for control animals or animals exposed to CuSO4, as determined by linear regression (lm function in R). n.s. = not significant, * p < 0.05, ** p < 0.01, *** p < 0.001. The Bonferroni method was used to adjust all p-values for multiple comparisons. Please click here to view a larger version of this figure.
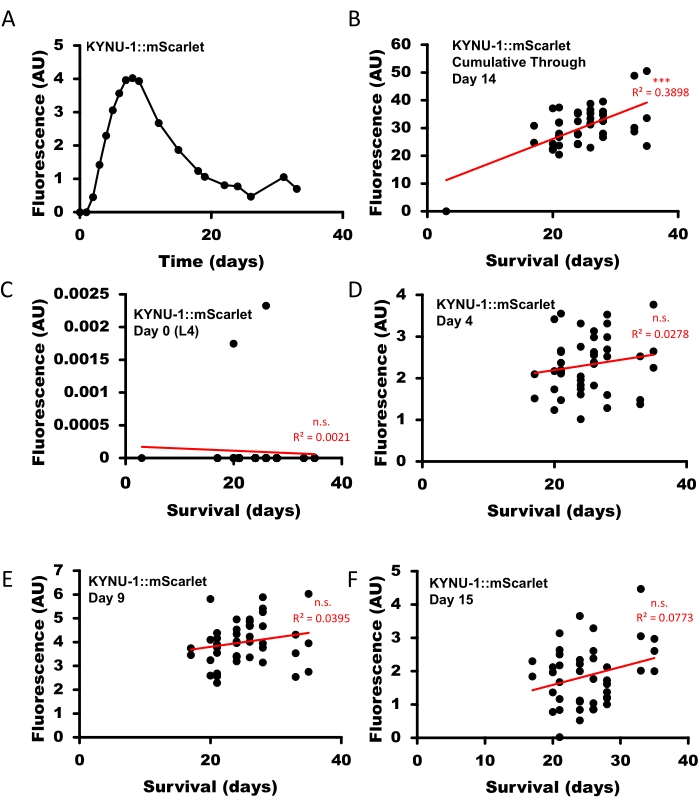
Figure 3: Dynamic quantification of KYNU-1 in individual C. elegans throughout the lifespan. C. elegans with mScarlet transgenically fused in-frame to the C-terminus of the kynu-1 gene (KYNU-1::mScarlet) were imaged using a fluorescent stereo microscope equipped with a monochrome camera every 1-4 days for 36 days until all animals had died. (A) KYNU-1, as measured by daily quantitation of KYNU-1::mScarlet fluorescence, shows a distinct age-dependent expression pattern across the lifespan (average value across worms shown at each age). (B) Cumulative KYNU-1::mScarlet expression through day 14 of adulthood correlates with lifespan in individual animals. KYNU1::mScarlet expression at the (C) L4 larval stage (day 0 of adulthood), (D) day 4 of adulthood, (E) day 9 of adulthood, and (F) day 15 of adulthood did not correlate with lifespan in individual animals. Correlations calculated by linear regression (Data Analysis Regression function in Microsoft Excel). n.s. = not significant, * p < 0.05, ** p < 0.01, *** p < 0.001. All p-values were adjusted for multiple comparisons using the Bonferroni method. Please click here to view a larger version of this figure.
Supplementary Figure 1: Single KYNU-1::mScarlet C. elegans imaged throughout lifespan under red channel. Single well of a prefabricated microtray containing a single C. elegans with mScarlet tagged KYNU-1 living on concentrated E. coli food source. The animal was first imaged at the L4 larval stage and then imaged at increments of 1-5 days for the remainder of its life. Please click here to download this File.
Supplementary Figure 2: Terasaki tray versus WorMotel background and artifact comparison under fluorescence microscopy. Wells of prefabricated microtrays (top) or molded PDMS WorMotels (bottom) were imaged under GFP (A), RFP (B), and DAPI (C) fluorescence channels using a fluorescent stereo microscope equipped with a monochrome camera. Images are shown in false color, and a heatmap is used to highlight points of saturation. Both culture systems have low-level backgrounds in all channels. The WorMotel plates are prone to sporadic high-signal artifacts, which are likely either microbubbles or dust or other particulate captured inadvertently during the PDMS molding process14,15. Please click here to download this File.
Supplemental File 1: Microtray_adapter_2021-10-20.STL. A stereolithography (STL) file for the 3D-printed insert on which the microtray is placed in the single-well plate. This file contains the full version with a solid bottom and is best suited for use in a system where the light source is above the plate. Please click here to download this File.
Supplemental File 2: Microtray_adapter_bottomless_2021-10-20.STL. A stereolithography (STL) file for the 3D-printed insert on which the microtray is placed in the single-well plate. This file contains the bottomless version and is best suited for use in a system where the light source is below the plate. Please click here to download this File.
