For up to 1 h, this procedure allows for easy measurement of intracellular changes in the fluorescence of monocarboxylate and glucose sensors. As shown in Figure 4, Laconic sensors in both glial cells and motor neurons respond to 1 mM lactate at a similar rate at the start of the pulse, but motor neurons reach a higher increase over the baseline during the 5 min pulse, as previously demonstrated17. This lactate concentration was chosen because it is comparable to the levels found in the hemolymph of third-instar larvae. Similarly, when VNC-expressing glucose sensors were exposed to 5 mM glucose (a value similar to that measured in the hemolymph by us and others19), the sensor's signal increased at a similar rate in glial cells and neurons. During the glucose pulse, however, the signal in neurons increases more than in glial cells. This is consistent with previous observations of similar FRET sensor-expressing preparations16.
This preparation can be used to conduct experiments on undescribed monocarboxylate and glucose transporters expressed in Drosophila glial cells or neurons20. Here we exemplify the use of RNA-interference to show that Chaski (Chk), which was previously known to transport monocarboxylates in heterologous cells, transports lactate in glial cells (Figure 5). Under nutritional restriction conditions, this transporter was described as playing an important role in animal physiology and behavior21. We found that in glial cells, knocking out Chk reduced lactate transport compared to control VNCs exposed to 1 mM lactate (Figure 5). Other putative transporters can be tested to see if they can transport metabolites in physiological and pathological conditions.
To address the study of metabolic changes associated with neuronal activity, we developed a simple method to increase neuronal activity without using technically complex procedures such as optogenetics, which can interfere with FRET sensor measurements (Figure 6). The isolated VNC was exposed to Picrotoxin (PTX), a known GABAA receptor antagonist previously used by us and others17,22. The concentration used increases the frequency of calcium oscillations in neurons (as measured by changes in GCaMP6f fluorescence) as well as significant changes in metabolically relevant molecules like glucose, lactate, and pyruvate17. Moreover, PTX-induced increases in neuronal activity result in a transitory drop in ATP levels in the soma of motor neurons. This phenomenon has previously been described in vertebrate models in which neuronal activity was increased via electric stimulation or by using GABAA receptor antagonists23,24. As a result, this model can be used to track metabolic changes in specific subsets of neurons and glial cells, addressing the need for energy-related transporters that contribute to ATP production during high neuronal activity.
Glucose and monocarboxylate transport are also important in tissues or cells other than the brain. We show here the visualization of monocarboxylate (lactate/pyruvate) and glucose uptake in fat bodies, a metabolically relevant tissue present in the larvae and the adult fly that has several key functions such as lipid storage and metabolization25. The Laconic sensor is well expressed in FB, as shown in Figure 7, where the lipid droplets can be seen as tiny black spheres within the cell. This isolated tissue adheres well to the poly-L-lysine-coated coverslips, allowing for a long-term cell imaging procedure that is also easy to analyze. We observed a significant increase in the fluorescence of the glucose sensor when FB was exposed to increasing glucose concentrations (approximately 5 mM), which is close to the glucose concentration found in the hemolymph. Further exposure to higher concentrations of glucose (10 mM) does not result in the expected increase in sensor fluorescence. Finally, in FB, we were able to measure lactate and pyruvate import (Figure 7C,D). In this case, increasing the lactate and pyruvate concentrations causes a proportional increase in Laconic and Pyronic fluorescence (ranging from 0.1 to 1 mM). Higher monocarboxylate concentrations result in signal saturation inside the cells.
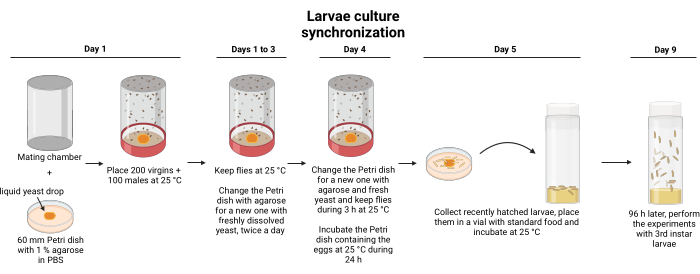
Figure 1: Synchronization of Drosophila larvae. The larval synchronization procedure is depicted graphically. Two hours before the process starts, 1% agarose plates must be prepared and changed twice daily until day 4. The hatching larvae must be collected with care. Please click here to view a larger version of this figure.
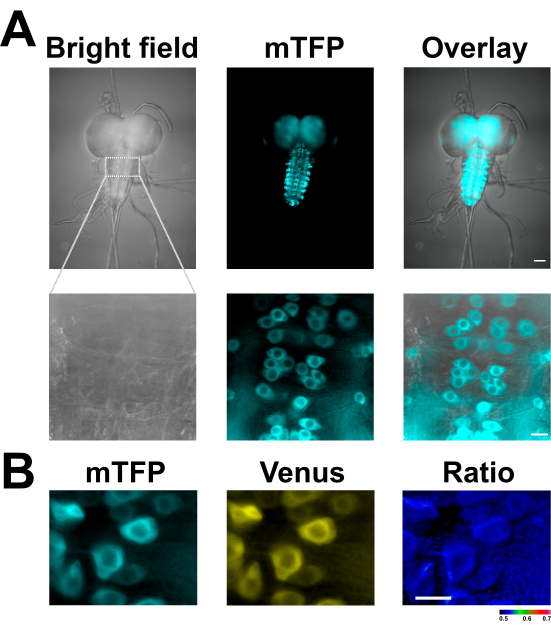
Figure 2: Isolated Drosophila larval brain expressing the lactate sensor Laconic. (A) A representative image of an isolated Drosophila third-instar larva's separated VNC adhered to a poly-L-lysine-coated coverslip and expressing the lactate sensor Laconic in motor neurons (OK6-GAL4). The images show a brightfield image of the VNC (left) and the sensor's mTFP fluorescence (middle panel) captured with a 20x water immersion objective. The images in the bottom panel are of the same VNC taken with a 40x water immersion objective. (B) Motor neurons from a VNC expressing the Laconic lactate sensor (OK6>Laconic) placed in a zero-lactate saline solution. The image depicts mTFP fluorescence (left), Venus fluorescence (center), and the ratio between the two fluorescence (right, produced using the Ratio Plus plugin of ImageJ software). Scale bars = 50 µm (A, top panel), 10 µm (A bottom panel, B). Abbreviations: VNC = ventral nerve cord; mTFP = monomeric teal fluorescent protein. Please click here to view a larger version of this figure.
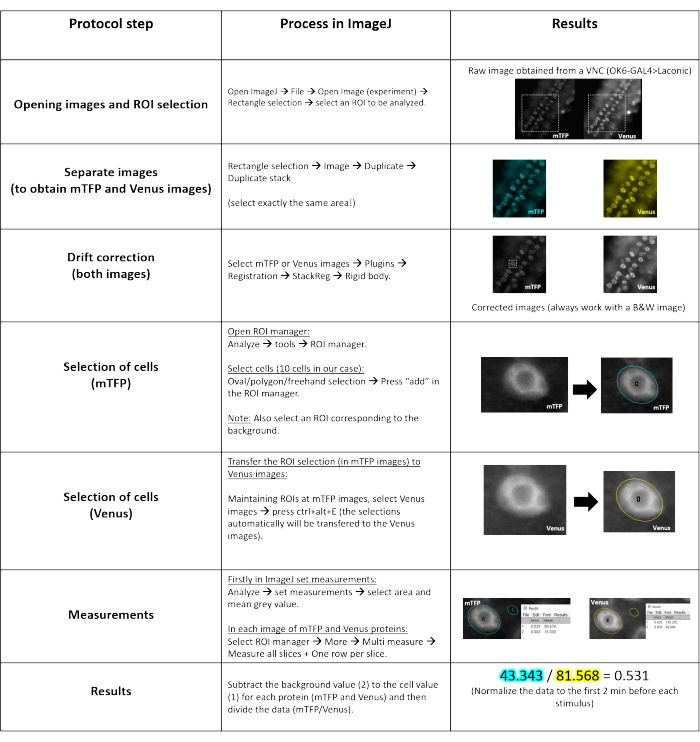
Figure 3: Step by step analyses of the images. A schematic illustration of the ImageJ software protocol for image analysis, displaying the process (first column), ImageJ commands (second column), and image processing outcomes (third column). The example starts with a raw image taken from a VNC expressing the Laconic lactate sensor in motor neurons (OK6-GAL4). Abbreviations: VNC = ventral nerve cord; mTFP = monomeric teal fluorescent protein. Please click here to view a larger version of this figure.
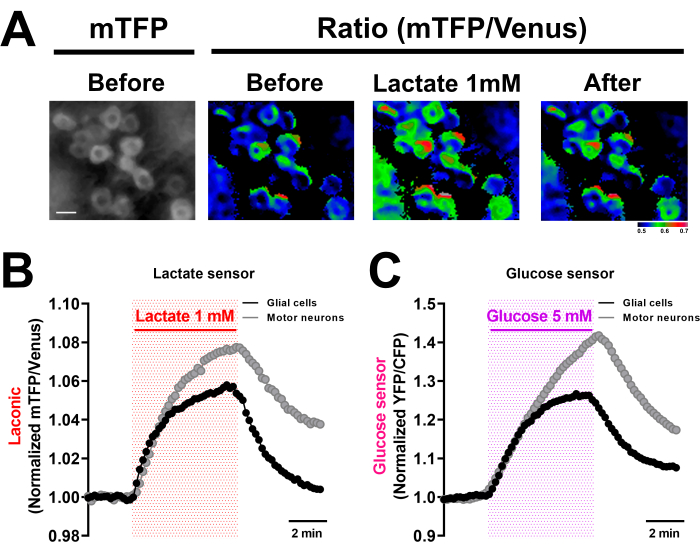
Figure 4: Monocarboxylate and glucose transport in neurons and glial cells of the Drosophila larval brain. (A) Representative images of VNC expressing the lactate sensor Laconic in motor neurons (OK6-Gal4>Laconic). Laconic fluorescence in motor neurons is seen before, during, and after a 5 min pulse of 1 mM of lactate. ImageJ's Ratio Plus plugin was used to create ratio images (mTFP/Venus). Scale bar = 10 µm. (B) Representative recordings of FRET signals from isolated VNCs expressing the Laconic lactate sensor in motor neurons (OK6-GAL4, grey lines) or glial cells (REPO-GAL4, black lines) exposed to 1 mM lactate for 5 min. The traces depict the adjusted FRET signals to the mean value collected 2 min before the lactate exposure. (C) Representative recordings of FRET signals from isolated VNCs expressing the glucose sensor FLII12Pglu700µδ6 in motor neurons (OK6-GAL4, grey lines) or glial cells (REPO-GAL4, black lines) exposed to 5 mM glucose for 5 min. The traces depict the adjusted FRET signals to the mean value collected two minutes before the glucose exposure. Abbreviations: VNC = ventral nerve cord; mTFP = monomeric teal fluorescent protein; FRET = Föster resonance energy transfer. Please click here to view a larger version of this figure.
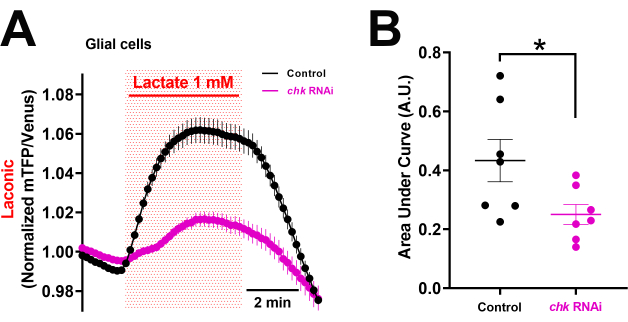
Figure 5: Lactate transport in glial cells of the Drosophila larval brain expressing Chaski RNAi. (A) Isolated VNC expressing the lactate sensor Laconic alone (genetic control, w1118, black line) or co-expressing an RNAi to knockdown Chaski (Chk) expression (chk-RNAi, magenta) were exposed to 1 mM Lactate for 5 min. For each condition, the traces represent the mean FRET value obtained from seven separate VNCs (70 cells per condition, normalized using the mean value obtained 2 min before being exposed to lactate). (B) The area under the curve in A is used to calculate intracellular lactate accumulation. For each condition, the values are the mean SE from 70 cells and seven independent experiments (control or chk-RNAi). The unpaired Student's t-test was used for statistical analysis (*p < 0.05). Abbreviations: VNC = ventral nerve cord; FRET = Föster resonance energy transfer. Please click here to view a larger version of this figure.
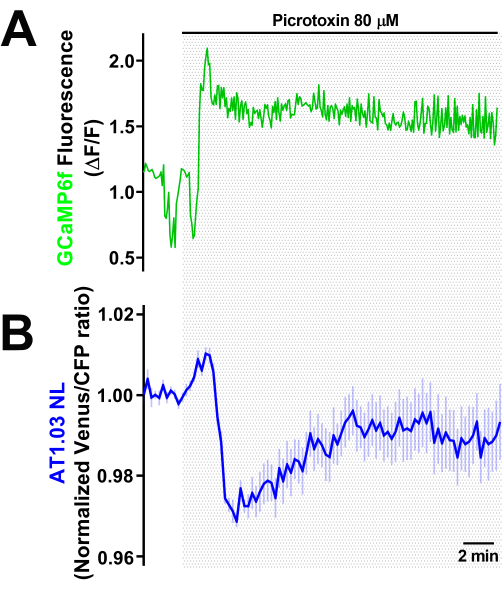
Figure 6: Changes in neuronal ATP levels induced by picrotoxin exposure. Recordings of the fluorescence captured from VNCs expressing the genetically encoded (A) calcium sensor GCaMP6f or (B, a different set of experiments) the ATP sensor AT1.03NL in motor neurons (OK6-GAL4) that were exposed to the GABAA receptor antagonist Picrotoxin (80 µM) during the time indicated by the line. The representative recording comes from a single motor neuron from a VNC (in A) or the mean ± SE from 10 cells from a single VNC (in B) normalized to the mean values obtained 2 min prior to PTX exposure. Abbreviations: VNC = ventral nerve cord; PTX = picrotoxin; SE = standard error; CFP = cyan fluorescent protein. Please click here to view a larger version of this figure.
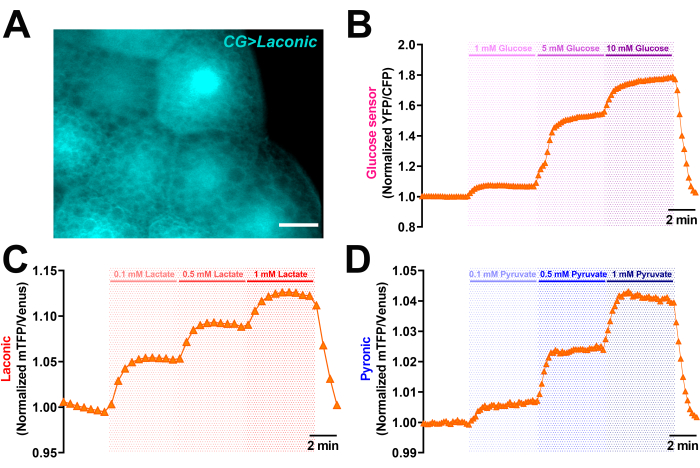
Figure 7: Transport of monocarboxylate and glucose in Drosophila fat bodies. (A) Image of the isolated fat body from third instar larvae expressing the lactate sensor Laconic (mTFP fluorescence). Scale bar = 10 µm. (B) Representative traces of the normalized signal captured in FB expressing the glucose sensor (FLII12Pglu700µδ6) exposed to glucose (1, 5, and 10 mM of glucose). Data were normalized to the first 2 min before glucose exposure. (C,D) Representative traces of the normalized signal obtained from FB expressing the lactate sensor (C) or the pyruvate sensor (D) exposed to 0.1-0.5-1 mM of lactate or pyruvate. The data were normalized to the first 2 min before lactate or pyruvate exposure. Abbreviations: FB = fat body; mTFP = monomeric teal fluorescent protein; YFP = yellow fluorescent protein; CFP = cyan fluorescent protein. Please click here to view a larger version of this figure.
