G蛋白偶联受体(GPCRs)构成所有细胞表面蛋白中最大和最多样化的家族之一。其在脊椎动物中存在,无脊椎动物,植物,酵母和粘菌,以及在原生动物和最早胚层后生动物表明GPCR是与信号转导1相连的最古老的分子之间。其天然配体的激活包括外部刺激包括肽,生物胺,增味剂,糖蛋白和光子2的广泛的多样性。正因为如此,这些受体 – 配体的信号系统涉及多种多样的生理过程。广泛的功能范围使它们非常适合的治疗药物,涵盖范围广泛的人类疾病的发展。约50-60%的电流药物靶点是通过G蛋白偶联受体3,4来表示。除了在制药行业的重视,G蛋白偶联受体也都在聚光灯下的发展新一代的种属特异性杀虫剂5,6和农药一般。因为很多G蛋白偶联受体的天然配体仍然不明,它们被归类为孤儿G蛋白偶联受体。这些受体的deorphanization将提高生物体的生理作用的认识,可能会发现假定目标为新药申请7。
由于基因组时代,反向药理学策略被广泛应用于对GPCR的8的deorphanization。该方法意味着一个孤儿受体被用来作为一个'钩'到'捞出'它从生物提取物或合成的化合物库激活配体。因此,感兴趣的G蛋白偶联受体被克隆,随后转染的细胞表达系统。在最常用的方法中,受体激活是通过测量变化的第二信使分子的浓度来决定9 </suP>。主要受体筛选分析依赖于钙敏感的生物发光蛋白( 如水母)10或荧光钙指标( 例如 ,荧光4)11。基于荧光的测定法,其中,受体表达细胞之前将配位体的筛选装有一个荧光钙指示剂,具有的优点是它们允许高通量筛选,因为它们易于使用,短的读取时间,并检查服务的灵活性多种孤儿受体上的单个板12。
在此,基于荧光的钙动员测定法中全面描述并通过果蝇短神经肽F(sNPF)受体的deorphanization过程示出。这neuropeptidergic信号系统最初是特征Mertens的等 。于2002年13与在中国仓鼠卵巢(CHO)细胞中进行的钙的生物发光测定法14和冯等人在2003年利用电生理测定爪蟾卵母细胞15。该sNPF信号系统的存在似乎只限于节肢动物门,它是牵连广泛的过程,包括摄食,生长,应激反应,运动和昼夜节律16的调节的门。
研究neuropeptidergic信号系统中的昆虫不仅可能导致对杀虫剂的发展新目标,但其运作的知识,也可以向尽可能多的信号系统已普遍非常保守的整个进化过程17其他生物推断。在过去的十年中,巨大的进步已经取得了昆虫神经肽G蛋白偶联受体的deorphanization过程。尽管有这些努力,只有少数的受体已被匹配到它们的同源配体,和序列信息的负荷为新的孤儿G蛋白偶联受体已成为可由于蓬勃发展的基因组学18。的中/高通量筛选的方法,例如,已被证明是一种广泛应用的技术9,18的基于荧光的钙动员测定法的可用性,因此非常宝贵的。
如这里所描述的基于荧光的钙动员测定法中的人胚肾293T(HEK293T)细胞系中进行,并使用荧光探针后,受体活化,以确定改变细胞内钙离子浓度。为了保证高表达与受体的翻译水平,Kozak共有序列19被添加到受体编码序列,它随后被克隆在表达载体中( 例如 ,pcDNA载体系列的哺乳动物细胞系)的5'末端。因为它是难以预料的内源性g的孤儿G蛋白偶联受体的基于序列信息蛋白偶联独自一人,第二信使分子( 如钙或营)正在受体激活后调制经常保持之前的配体识别未知。为了解决这个问题,对G q系列( 如鼠Gα15或人类Gα16 [这里使用])或嵌合G蛋白( 例如 ,Gαqi5)的混杂G蛋白与G蛋白偶联受体大部分互动和诱导钙的释放可共表达20,21,22。根据该配体与其受体的结合,GPCR的发生,导致特定的细胞内途径的激活的构象变化。鸟苷二磷酸(GDP)的分子,休息条件的Gα亚基16的约束下,将通过三磷酸鸟苷(GTP)分子所取代。这激起了异三聚体G蛋白的解离在一个Gα16和Gβγ亚单位。该Gα亚基16激活磷脂酶C&#946; (PLCβ),这反过来水解与膜结合的磷脂酰肌醇二磷酸(PIP 2)产生二酰基甘油(DAG)和三磷酸肌醇(IP 3)。 IP 3会扩散到整个细胞质和激活的IP 3依赖性钙通道存在于内质网,其诱导钙释放到细胞质中的膜。
在受体活化的钙释放发生在几秒钟内,并且可以通过筛选试验之前加载的细胞用钙敏感的染料,如荧光4乙酰氧基甲基(AM)11来检测。对AM酯基使荧光穿过细胞膜,并通过细胞质酯酶的细胞一旦内切割掉。因此,在荧光染料的负电荷被屏蔽,防止它扩散出细胞,并允许它与钙离子进行交互。荧光信号Of荧光4是微不足道的细胞中只含有在纳摩尔范围内钙离子浓度静息状态下。然而,当钙被释放后受体激活,信号可以增加浓度依赖性地超过了100倍,因此保证大的信号 – 噪声比。荧光4也表现出大动态范围的报告[钙]周围的K 345 nM的D(钙),使其适用于测量各种细胞的生理有关钙的变化。的Fluo-4的激发发生在488nm和发射的荧光的测量是在525纳米11。象的荧光成像板读数器(FLIPR)23,可以在Novostar或FlexStation(台装置)12是荧光计中/高通量系统,允许同时进行化合物加成并且在受体活化的Fluo-4的信号的检测对于每个井在测定板。这里所描述的钙动员检测依赖于站上设备96孔微孔板的系统。
本的SoftMax Pro软件(软件),用于操作该站装置,以及对数据进行分析。该程序将立即显示的结果为在96 – 孔格式的图形。多个孔可以同时被选择以这些油井的结果比较在相同的曲线图。孔中的每个列的相对荧光单位(RFU)的值是同时测量的两分钟内,开始加入的化合物的各孔中之前和之后的受体激活的荧光信号的测量后继续进行。通常,激动剂曲线的趋势对齐基线直到活化化合物加入到细胞中,从而导致荧光信号的快速增加。的峰值高度相关,在该孔的最终激动剂浓度。高峰期后,荧光信号缓慢下降对基线水平。俄罗斯足协测量CAn为转换成浓度-响应曲线,以确定配体的EC 50值(半数最大有效浓度)。在一般情况下,至少三个独立的屏幕,每一个都包括的浓度系列三个副本,应执行以构成可靠的浓度 – 反应曲线。
建议将包括在实验设计中的几个阳性和阴性对照。首先,一个转染对照, 即用已知的配体的受体的实施,应进行测试。这使得验证转染试剂是否运作。掺入 的对照实验用的激动剂的细胞系和阴性对照( 例如 ,洗涤缓冲液)的内源性受体,也推荐以监测细胞的健康和生存能力,并排除该洗涤缓冲液被污染的可能性这可能引起自动fluore一个因素气味的反应。常用的激动剂是从蛋白酶激活受体-1(PAR 1),它作为一个PAR 1选择性激动剂,或氯化氨甲酰胆碱,激活乙酰胆碱受体的肽。转染空载体的表达也应该进行测试,以排除活性化合物与细胞的内源性受体相互作用。可能需要不同的信令系统在下面的协议描述的几个参数优化。完整的基于荧光的钙动员测定法的概略图是描绘在图1中 。
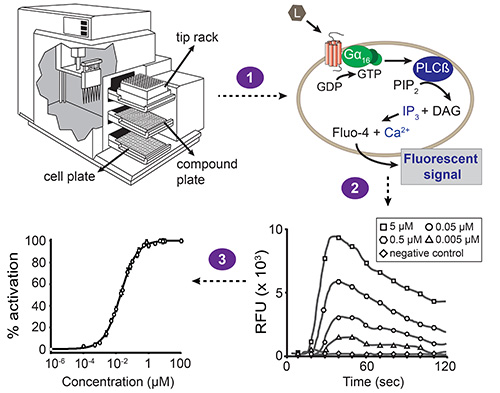
基于荧光的钙动员检测的图1,总体方案。自动化液体处理和同步荧光测量与台执行设备酶标仪,由软件驱动。站设备包含三个抽屉:一个用于使单元板,复合板和吸头机架。积聚在移液器转移化合物从化合物板的一列,以使单元板(步骤1)的相应列中。细胞板的每个孔中包含已被共转染与所关注的GPCR和混杂Gα亚基16的HEK293T细胞的单层。当一种化合物激活受体,使Gα16结合的GDP被替换为GTP。的Gα亚基16随后解离的Gβγ复合物,并激活磷脂酶Cβ(PLCβ),这反过来水解磷脂酰肌醇二磷酸(PIP 2)产生二酰基甘油(DAG)和三磷酸肌醇(IP 3)。 IP 3激活的IP 3依赖性钙通道存在于内质网的膜,诱导钙整数的释放Ø细胞质中。钙与的Fluo-4(与该细胞为化合物加成之前加载)的相互作用导致的荧光信号(步骤2)。软件给出的结果作为相对荧光单元中的时间函数(RFU)的值,以及峰值高度相关与配位体浓度的浓度依赖性。这些数据可以被转换成浓度-响应曲线,以确定配体-受体对(步骤3)的EC 50值。
