Renale epitel iontransport ligger til grund organismal iono- og osmoregulering. Drosophila melanogaster Malpighian (renal) tubulus tilbyder en enestående mulighed for at studere de molekylære mekanismer i epitelial ion transport. Dette skyldes kombinationen af de stærke genetik Drosophila, parret med tilgængeligheden af sine nyretubuli til fysiologisk undersøgelse. Ramsay assay, opkaldt efter den undersøger, som banebrydende teknik 1, måler væskesekretion satser fra isolerede Malpighian tubuli, og blev etableret i Drosophila i 1994 af Dow og kolleger 2. Dette banede vejen for yderligere undersøgelser ved hjælp af Drosophila genetiske værktøjer, såsom GAL4-UAS systemet 3,4, for at definere celle-specifikke signalveje regulerer væske sekretion. Et eksempel omfatter calcium signalering som respons på et peptidhormon 5, blandt mange andre 6,7.
ve_content "> en kombination af genetiske teknikker og klassiske fysiologiske undersøgelse har vist, at urin generation i fluen sker gennem sekretion af et kaliumchlorid-rige væske fra hovedsegmentet af tubuli. Dette sker via den parallelle transepitel sekretion af kationer, primært K + men også Na +, gennem den primære celle og Cl -. sekretion gennem stjerneformet cellen 8-12 Evnen til separat at måle transepiteliale K + og Na + flux giver en mere detaljeret karakterisering af transportmekanismer end måling af væske sekretion alene. For eksempel i ustimulerede Drosophila tubuli, Na + / K + -ATPase-inhibitoren ouabain har ingen effekt på fluid sekretion 2, selv når dets optagelse i celler vigtigste inhiberes af organisk aniontransporter inhibitor taurocholat 13. Imidlertid Linton og O'Donnell viste, at ouabain depolarisererden basolaterale membran potentiale, og øger Na + flux 9. Som vist i Repræsentative resultater, vi gentaget disse resultater, og viste, at K + flux er samtidig faldet 14; den øgede Na + flux og faldt K + flux har modsatrettede virkninger på fluid sekretion, hvilket resulterer i nogen nettoændring i sekretion. Der er således to beslutninger til "ouabain paradoks", dvs. den oprindelige iagttagelse, at ouabain har nogen virkning på fluid sekretion i Drosophila tubulus:. Dels i stimulerede tubuli, virkningen af ouabain på fluid sekretion fremgår ikke på grund af dens optagelse af organisk aniontransporter 13; og for det andet i ustimulerede tubuli, ouabain har modstående virkninger på transepitelialt Na + og K + flux, hvilket resulterer i nogen nettoændring i fluid sekretion (se Repræsentative resultater og ref. 9). Derfor er den primære rolle af Na + / K + ATPase i ustimulerede tubuli er at sænke intracellulære Na + koncentration til at frembringe en positiv koncentrationsgradient for Na + -coupled transportprocesser på tværs af basolaterale membran. Faktisk ved separat måling Na + og K + fluxe, vi påvist, at tubuli mangler fluen natrium-kalium-2-chlorid cotransportøren (NKCC) er faldet transepitelial K + flux, uden yderligere fald efter ouabain Desuden, og ingen ændring i transepitel Na + flux 14. Disse resultater støttede vores konklusion, at Na + ind i cellen gennem NKCC recirkuleres gennem Na + / K + -ATPase. I et andet eksempel Ianowski et al. Observerede, at en sænkning bad K + koncentration fra 10 mM til 6 mM reduceret transepitheliale K + flux og forøget transepitel Na + flux i tubuli fra Rhodnius prolixus, med nogen nettoændring i fluid sekretion <sup> 15. Forskellige virkninger på Na + flux og K + flux på tværs af larver tubuli er også blevet observeret i Drosophila tubuli som reaktion på varierende salt kost 16 og i to myg arter som reaktion på opdræt saltholdighed 17.Den største udfordring i målingen af transepitelial ion flux i Ramsay assay forberedelse er bestemmelsen af ion koncentrationer inden det udskilte væske. Denne udfordring er blevet mødt med varierende løsninger, herunder flamme photometery 18, anvendelse af radioaktive ioner 19, og elektron sonde bølgelængde dispersivt spektroskopi 20. Disse teknikker kræver overførsel af den udskilte væske fald til et instrument til måling af ionkoncentrationer. Da mængden af væske, der udskilles af den ustimulerede Drosophila tubulus er lille, typisk ~ 0,5 nl / min, udgør det en teknisk udfordring, og også indfører fejl, hvis nogle af de udskilte væsketabt ved overførsel. I modsætning hertil anvendelsen af ionspecifikke elektroder tillader måling af ion aktivitet (hvorfra der kan beregnes ionkoncentration) in situ. Den nuværende protokol blev tilpasset fra den, der anvendes af Maddrell og kolleger til at måle transepitel K + flux på tværs af Rhodnius tubulus hjælp valinomycin som K + ionofor 21 og beskriver også anvendelsen af en 4-tert -butylcalix [4] aren-tetraeddikesyre tetraethylester-baserede Na + -specifik ion-specifik elektrode kendetegnet ved Messerli et. al. 22. Ion-specifikke elektroder er også blevet anvendt til at måle ion koncentrationer i væske, der udskilles af Malpighian tubuli i Ramsay assay voksne 9,23 og larver 16 Drosophila melanogaster, New Zealand Alpine Weta (Hemideina Maori) 24 og myg 17.
Her beskriver vi i detaljer brugen af Ramsay somsige at måle fluid sekretionshastigheder i Malpighian tubuli fra Drosophila melanogaster, samt anvendelsen af ionspecifikke elektroder til at bestemme koncentrationerne af K + og Na + i den udskilte væske og dermed beregningen af transepitelialt ionstrømme. En oversigt over assayet er tilvejebragt i figur 1.
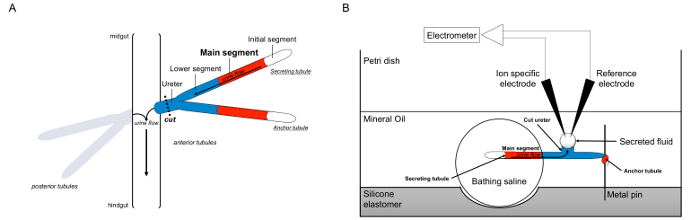
Figur 1. Skematisk af Malpighian tubuli og Ramsay-analysen med brug af Ion-specifikke elektroder til måling ionkoncentrationer. Denne figur illustrerer opsætningen for Ramsay analysen. (A) Hver flue har fire tubuli, et par anteriore tubuli og et par posteriore tubuli, der flyder i bughulen omgivet af hæmolymfe. I hvert par, de to tubuli deltage i urinlederen, som derefter tømmer urinen ved krydset af mellemtarmen og hindgut. Tubuliene er blinde ender. Urin frembringes af fluid-udskillende hovedsegmentet (vist med rødt) og strømmer mod ureter og ud i tarmen. Efter dissektion bliver tubulus parret adskilt fra tarmen ved transecting ureter. (B) Parret af tubuli overføres derefter til en dråbe saltvand badning inden for en brønd i assay skålen. En af de to tubuli, der er nævnt her som "anker tubulus," er viklet omkring en metalstang og er inert. Den anden tubulus er udskillende tubulus. Den indledende segment (som ikke udskiller væske) og vigtigste segment af secernerer tubulus forbliver inden for dråbe badning saltvand. Ioner og vand træk fra badning saltvand og ind i tubulus lumen de vigtigste segment, og derefter bevæge sig mod ureter, som ville forekomme in vivo. Den nederste segment (blå) er uden for badning saltvand og derfor inaktivt. Da ureter skæres, fremgår det secernerede fluid som en dråbe fra den afskårne ende af ureter. Than udskilte væske dråbe forstørrer, efterhånden som sekretion fortsætter, og dens diameter er målt ved anvendelse af en okulær mikrometer. Et lag af mineralolie forhindrer fordampning af det udskilte væske. Reference- og ion specifikke elektroder måler ion koncentrationen af det udskilte væske. Klik her for at se en større version af dette tal.
