신장 상피 이온 수송은 유기체의 iono- 및 삼투의 기초가. 초파리 melanogaster의 Malpighian (신장) 세관 상피 이온 수송의 분자 메커니즘을 연구 할 수있는 최고의 기회를 제공합니다. 이것은 생리 학적 연구에의 신장 세뇨관의 접근성과 짝을 초파리의 강력한 유전학의 조합 때문이다. 기술 1을 개척 연구자의 이름을 따서 명명 램지 분석, 격리 Malpighian 세관에서 유체 분비 속도를 측정하고, 다우와 동료 2에 의해 1994 년에 초파리 년에 설립되었습니다. 이것은 유체 분비 조절 셀 고유 신호 전달 경로를 정의하는, 초파리 등을 GAL4-UAS 시스템 3,4- 유전 도구를 사용하여 추가 연구를위한 기반을 마련. 예는 많은 다른 6,7 사이에 펩티드 호르몬 (5)에 응답하여 칼슘 신호를 포함한다.
비행 세뇨관의 주요 부분에서 염화칼륨이 풍부한 액체의 분비를 통해 발생에 ve_content "> 유전자 기술 및 고전 생리 학적 연구의 조합 뇨 생성을 도시하고있다.이 주로 양이온 평행 transepithelial 분비 통해 이루어 K는 +뿐만 아니라 나트륨 +는 주체 세포, 및 CL를 통해 -. 성상 세포 8-12 내지 분비 별도로 transepithelial K + 및 나트륨 + 플럭스를 측정하는 능력은 유체 분비의 측정보다 반송기구의 상세한 특성화를 허용 홀로. 예를 들어, 자극되지 초파리 세뇨관에서, 나 +가 / K + -ATPase 억제제 인 ouabain 유체 분비 2에 영향을주지 주체 세포 내로의 흡수량은 유기 음이온 수송 체 억제제 타우로 콜산 (13)에 의해 억제 된 경우에도. 그러나, 린톤 및 오도넬은 ouabain과는 탈분극 것을 보여 주었다기저 막 잠재력, 그리고 나 + 플럭스 (9)을 증가시킨다. 대표 결과에 도시 된 바와 같이, 우리는 이러한 발견을 복제하고, 14 병용 감소 K + 플럭스가 있음을 보여 주었다; 증가 나 + 플럭스 및 K 감소 + 플럭스 분비없는 순 변화의 결과로 유체 분비에 반대하는 효과를 가지고있다. . 따라서, "ouabain과 모순", 즉 두 가지 해결책이 존재하며, 초파리 세뇨관 유체 분비에 영향을주지 ouabain과 초기 관찰 : 제 자극 세뇨관에서 유체 분비 ouabain과 효과는 명백하지 않다 인한 유기 음이온 수송 (13)에 의해 그 흡수; 둘째, 자극되지 세관에서, ouabain과는 (대표 결과와 심판을 참조하십시오. 9) 유체 분비없는 순 변화의 결과로, transepithelial 나 +와 K + 플럭스에 영향을 반대하고있다. 따라서, 나 +의 주요 역할 / K + -ATP비자극 세뇨관에서 ASE는 나트륨 +는 기저 외측 막에 걸쳐 전송 프로세스 -coupled위한 유리한 농도 기울기를 생성하는 세포 나 + 농도를 낮출 수있다. 실제로, 별도로 나 +와 K + 플럭스를 측정하여, 우리는 세관이 비행 나트륨 – 칼륨 – 2 – 염화 송체 (NKCC)를 부족하지 ouabain과 첨가 한 후 더 이상의 감소, transepithelial K + 플럭스 감소했으며, transepithelial의 변화가 있음을 보여 주었다 나 + 14 플럭스. 이러한 연구 결과는 NKCC를 통해 세포를 입력 나 +가 나 + / K + -ATPase를 통해 재활용 우리의 결론을지지했다. 다른 예에서, Ianowski는 외. mM의 10 mM 내지 6 욕 K + 농도를 낮추면 유체 분비없는 순 변화와 Rhodnius의 prolixus에서 세뇨관에서 transepithelial의 K + 플럭스 증가 transepithelial 나 + 플럭스 감소 관찰 <sup> 15. 애벌레 세관에서 나 + 플럭스 및 K + 플럭스에 차등 효과는 염분 (17) 양육에 대한 응답으로 다양한 소금 다이어트 (16)와 두 모기 종에 대한 응답으로 초파리 세관에서 관찰되었다.램지 분석 준비 transepithelial 이온 플럭스의 측정에서 가장 큰 도전은 분비 된 유체 내 이온 농도의 판정이다. 이 문제는 불꽃 photometery (18), 방사성 이온 (19)의 사용, 전자 프로브 파장 분산 분광기 (20)를 포함하여 다양한 솔루션을 만났다되었습니다. 이러한 기술은 이온 농도의 측정을위한 도구로 분비되는 액체 방울의 전송을 필요로한다. 비자극 초파리 세뇨관 분비 유체의 부피가 작기 때문에, 일반적으로 ~ 0.5 NL / 분, 이것은 기술적 도전을 제기하고 분비 된 유체의 일부가 오류가있는 경우에도 소개이전에 잃었다. 대조적으로, 특정 이온 전극의 사용은 인 시츄 (이온 농도가 계산 될 수있는) 이온 활성의 측정을 허용한다. 현재 프로토콜은 K + 이온 운반체 (21)로 발리 노마 이신 (Valinomycin) 사용 Rhodnius의 세뇨관 걸쳐 transepithelial의 K + 플럭스를 측정하도록 Maddrell 동료가 사용하는 각색 및 또한 4- 급의 -butylcalix [4] 아렌 – 테트라 아세트산의 사용을 설명했다 테트라 에틸 에스테르 계 나은 등 Messerli 특징으로 특이 이온 특정 전극을 +. 등. (22). 이온 특정 전극 성인 9,23과 애벌레 (16) 초파리에서 램지 분석에 Malpighian 세관에 의해 분비되는 유체의 이온 농도를 측정하는 데 사용 된, 뉴질랜드 알파인 웨타 (Hemideina 마오리어) (24) 모기 (17).
여기서는 상세히 램지의 사용 등을 서술유체 분비 초파리 melanogaster의 발 Malpighian 세뇨관의 속도뿐만 아니라 분비 유체 내 K + 및 나트륨 +의 농도를 결정하는 이온 고유의 전극을 사용하므로 transepithelial 이온 플럭스의 계산을 측정하기 말한다. 분석의 개요는도 1에 제공된다.
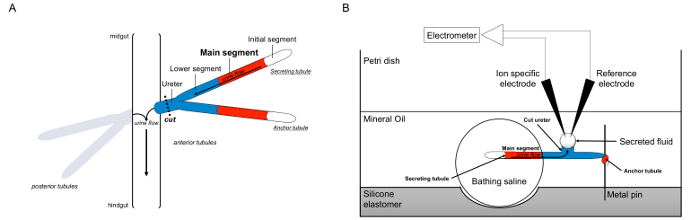
Malpighian 세관의 그림 1. 도식과 이온 농도를 측정하기 위해 이온 특정 전극의 사용과 램지 분석. 이 그림은 램지 분석에 대한 설정을 보여줍니다. (A) 각 플라이는 네 세뇨관, 전방 세뇨관의 쌍 및 후방 세뇨관의 쌍에 의해 둘러싸인 체액 복강 그 플로트있다. 각 쌍에서 두 세관은 중장 hindgu과의 교차점에 뇨를 비운다 요관에서 합류티. 세관은 블라인드 종료됩니다. 소변 (빨간색으로 표시) 유체 분비 주요 세그먼트에 의해 발생하고, 장에 요관으로 밖으로 흐른다. 절개 후 세관 쌍 요관을 직감에 병행하여 해리된다. (B)은 세뇨관의 쌍은 분석 접시의 웰 내에서 입욕 식염수 방울로 전달된다. 여기에 언급 된 두 세뇨관의 일 "앵커 세관은,"금속 핀 감싸 불활성이다. 다른 세관은 분비의 가느 다란 관이다. 초기 세그먼트 (유체 분비하지 않는)과 분비 세관의 주요 부분은 목욕 식염수의 액적 내에 남아있다. 생체 내에서 발생하는 것에 이온과 목욕 식염수에서 주 세그먼트의 세관 루멘에 물 이동, 그리고, 요관으로 이동합니다. 낮은 세그먼트 (파란색) 목욕 식염수 때문에 불활성 밖에 있습니다. 요관이 절단되기 때문에, 유체는 분비 요관의 절단 단부로부터 액적으로 나온다. 티그 분비는 계속 유체 액적 시간 경과 확대 분비, 그 직경은 접안 마이크로 미터를 사용하여 측정된다. 미네랄 오일 층은 분비 된 유체의 증발을 방지한다. 참조 및 이온 특정 전극이 분비 유체의 이온 농도를 측정한다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
