Transporte iónico epitelial renal subyace ionosfera del organismo y la osmorregulación. La Drosophila melanogaster Malpighi (renal) túbulo ofrece una oportunidad sin precedentes para estudiar los mecanismos moleculares de transporte iónico epitelial. Esto es debido a la combinación de las poderosas genética de Drosophila, emparejado con la accesibilidad de sus túbulos renales a estudio fisiológico. El ensayo de Ramsay, el nombre del investigador que fue pionero en la técnica 1, mide las tasas de secreción de líquido de los túbulos de Malpighi aislados, y se constituyó en Drosophila en 1994 por Dow y colegas 2. Esto allanó el camino para más estudios utilizando Drosophila herramientas genéticas, tales como el sistema GAL4-UAS 3,4, para definir las vías de señalización de células específicas que regulan la secreción de fluidos. Un ejemplo incluye la señalización del calcio en respuesta a una hormona peptídica 5, entre muchos otros 6,7.
ve_content "> Una combinación de técnicas genéticas y estudio fisiológico clásica ha demostrado que la generación de la orina en el se produce la marcha a través de la secreción de un cloruro de potasio rica en líquido de la principal segmento del túbulo. Esto ocurre a través de la secreción transepitelial paralelo de cationes, principalmente K + Na +, sino también, a través de la célula principal, y Cl -. a través de la secreción de las células estrelladas 8-12 La capacidad de medir por separado transepiteliales flujos de K + y Na + permite una caracterización más detallada de los mecanismos de transporte que la medición de la secreción de fluidos solo. Por ejemplo, en los túbulos de Drosophila no estimuladas, el Na + / K + -ATPasa ouabaína no tiene ningún efecto sobre la secreción de fluido 2, incluso cuando su captación en las células principales se inhibe por el transportador de aniones orgánicos inhibidor de taurocolato 13. Sin embargo, Linton y O'Donnell mostró que ouabaína despolarizael potencial de membrana basolateral, y aumenta Na + flujo 9. Como se muestra en los resultados representativos, replicamos estos hallazgos, y demostramos que K + flujo es concomitante disminución de 14; el aumento del flujo de Na + y la disminución de K + flujo tener efectos opuestos sobre la secreción de fluidos, lo que resulta en ningún cambio neto en la secreción. Por lo tanto, hay dos resoluciones a la "paradoja ouabaína", es decir, la observación inicial de que ouabaína no tiene ningún efecto sobre la secreción de fluido en el túbulo Drosophila:. Primero, en los túbulos estimulados, el efecto de ouabaína sobre la secreción de fluido no es evidente debido a la su absorción por el transportador de aniones orgánicos 13; y en segundo lugar, en los túbulos no estimuladas, ouabaina ha oponerse efectos sobre transepitelial Na + y K + flujo, lo que resulta en ningún cambio neto en la secreción de fluidos (ver resultados representativos y ref. 9). Por lo tanto, la función principal de la Na + / K + -ATPase en los túbulos no estimuladas es disminuir la concentración intracelular de Na + para generar un gradiente de concentración favorable para Na + -junto procesos de transporte a través de la membrana basolateral. En efecto, mediante la medición por separado de Na + y K + fundentes, hemos demostrado que los túbulos que carece del cotransportador sodio-potasio-2-cloruro de mosca (NKCC) han disminuido K transepitelial + fundente, sin mayor descenso después de la adición ouabaina, y ningún cambio en transepitelial Na + Flujo 14. Estos resultados apoyan nuestra conclusión de que Na + entrar en la célula a través de la NKCC se recicla a través de la Na + / K + -ATPasa. En otro ejemplo, Ianowski et al. Observaron que la reducción de baño de K + concentración de 10 mM a 6 mM disminuyó transepitelial K + y el aumento de flujo transepitelial Na + en los túbulos flujo de Rhodnius prolixus, sin cambio neto en la secreción de fluidos <sup> 15. Efectos diferenciales sobre Na + y K + flujo flujo a través de los túbulos de larvas se han observado también en los túbulos de Drosophila en respuesta a diferentes dietas de sal 16 y en dos especies de mosquitos en respuesta a la cría de salinidad 17.El mayor reto en la medición del flujo de iones transepitelial en la preparación de ensayo Ramsay es la determinación de las concentraciones de iones dentro del fluido secretado. Este reto se ha cumplido con soluciones diferentes, incluyendo photometery llama 18, el uso de iones radiactivos 19, y el electrón sonda de longitud de onda de la espectroscopía dispersiva 20. Estas técnicas requieren la transferencia de la gota de líquido secretado a un instrumento para la medición de concentraciones de iones. Dado que el volumen de líquido secretado por el túbulo Drosophila no estimulada es pequeño, típicamente ~ 0,5 nl / min, esto plantea un reto técnico y también introduce error si algo del fluido es secretadaperdido en la transferencia. En contraste, el uso de electrodos de ión específico permite la medición de la actividad de iones (a partir del cual se puede calcular la concentración de iones) in situ. El protocolo actual fue adaptado de la utilizada por Maddrell y sus colegas para medir transepitelial K + flujo a través de los túbulos Rhodnius usando valinomicina como el K + ionóforo 21, y también describe el uso de un -butylcalix 4- terc [4] areno-ácido tetraacético Na base de éster de tetraetilo + específico de electrodo específico de iones caracterizado por Messerli et. al., 22. Ion-electrodos específicos también se han utilizado para medir las concentraciones de iones en el fluido secretado por los túbulos de Malpighi en el ensayo de Ramsay en adulto y larval 9,23 16 Drosophila melanogaster, la Nueva Zelanda Alpine Weta (Hemideina maori) 24 y 17 en los mosquitos.
Aquí se describe en detalle el uso del Ramsay comodecir para medir las tasas de secreción de fluidos en los túbulos de Malpighi de Drosophila melanogaster, así como el uso de electrodos de ión específico para determinar las concentraciones de K + y Na + en el fluido secretado y por lo tanto el cálculo de los flujos de iones transepitelial. Una visión general del ensayo se proporciona en la Figura 1.
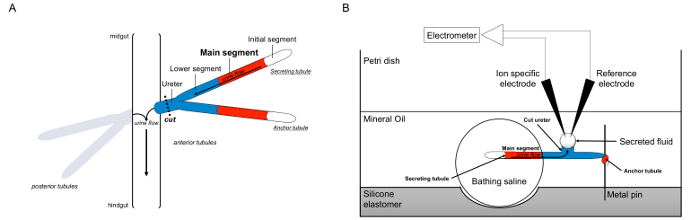
Figura 1. Esquema del Malpighi túbulos y el ensayo de Ramsay con uso de iones específicos electrodos para medir concentraciones de iones. Esta figura ilustra la configuración para el ensayo de Ramsay. (A) Cada mosca tiene cuatro túbulos, un par de túbulos anterior y un par de túbulos posteriores, que flotan en la cavidad abdominal rodeado de hemolinfa. En cada par, los dos túbulos se unen en el uréter, que luego se vacía la orina en la unión del intestino medio y hindgut. Los túbulos son ciego terminado. La orina se genera por el segmento principal de fluido que secretan (mostrado en rojo), y fluye hacia el uréter y hacia fuera en el intestino. Después de la disección, el par túbulo se disocia desde el intestino por transección el uréter. (B) El par de túbulos se transfiere a continuación en una gota de solución salina de baño dentro de un pocillo de la placa de ensayo. Uno de los dos túbulos, que se refiere aquí como el "túbulo ancla," se envuelve alrededor de un pasador de metal y es inerte. El otro es el túbulo túbulo secretor. El segmento inicial (que no secretar fluido) y el segmento principal de los túbulos secretores se mantienen dentro de la gota de solución salina de baño. Iones y movimiento de agua de la solución salina de baño y en el lumen de los túbulos del segmento principal, y luego se mueven hacia el uréter, como ocurriría en vivo. El segmento inferior (azul) está fuera de la solución salina de baño y por lo tanto inerte. Desde el uréter se corta, el fluido secretado emerge como una gotita desde el extremo cortado del uréter. Tél secretada gotita de fluido aumenta con el tiempo como la secreción continúa, y su diámetro se mide utilizando un micrómetro ocular. Una capa de aceite mineral evita la evaporación del fluido secretado. Los iones de referencia y electrodos específicos miden la concentración de iones del líquido secretado. Por favor haga clic aquí para ver una versión más grande de esta figura.
