الانصهار الغشاء هو عملية بيولوجية عالمية المطلوبة للاتجار داخل الخلايا من الدهون والبروتينات، إفراز، والتسميد، والتنمية، ويلفها دخول الفيروس إلى الكائنات المضيفة 1-3. بالنسبة لمعظم تفاعلات الاندماج داخل الخلايا بما في ذلك الإفراج عن الهرمونات والناقلات العصبية عبر إيماس، يتم توفير الطاقة لدمج اثنين من طبقات ثنائية الدهون عن طريق تشكيل لحزمة أربعة الحلزون بين (الفخ) البروتينات وما شابه ذلك للذوبان N-ethylmaleimide تراعي بروتين عامل مرفق مستقبلات، الراسية في الحويصلة (الخامس الفخ) والغشاء الهدف (تي الفخ) 4 على التوالي. متشابك الحويصلة إيماس هو تفاعل الاندماج أكثر ينظم بإحكام ويحدث خلال ميلي ثانية واحدة بعد وصول ل1،4،5 المحتملين عمل. المسام الانصهار، الاتصال الأولي بين اثنين من الصمامات مقصورات، يمكن أن تومض مفتوحة ومغلقة عدة مرات قبل إعادة طبع أو توسيع بصورة لا رجعة فيها 5-7. النتائج السابقةفي عابرة، "قبلة وتشغيل" الانصهار، في حين يؤدي الأخيرة إلى الانصهار الكامل. العوامل التي تحكم التوازن بين هذين الوضعين الانصهار وآليات تنظيم المسام الخفقان ليست مفهومة جيدا 5،8.
مطلوبة البروتينات كمين لإيماس. وألغت متشابك الانصهار الحويصلة على الانقسام من الافخاخ التي كتبها أعصاب 9. وأظهرت التجارب الانصهار السائبة باستخدام الحويصلات unilamellar الصغيرة (سيارات الدفع الرباعي) أن الافخاخ ليست مطلوبة فقط، ولكن أيضا ما يكفي لدفع الغشاء الانصهار 10. في هذا الاختبار بكميات كبيرة، وسيارات الدفع الرباعي أعادت مع الخامس الافخاخ (الخامس SUV) ومخدر مع الفوسفورية الفلورسنت (N – (7-نيترو-2-1،3-benzoxadiazol-4-YL) -phosphoethanolamine (NBD-PE) و ( N – (lissamine رودامين B السلفونيل) -phosphoethanolamine (LR-PE) ويخلط مع حويصلات غير المسماة التي تحتوي على تي الافخاخ (ر-SUV) في البداية مضان بنك دبي الوطني-PE في الخامس سيارات الدفع الرباعي ومروي التي كتبها فورستر نقل الطاقة الرنين (الحنق ) إلى LR-PE. كما مختبرالهنداوي الخامس سيارات الدفع الرباعي تلتحم مع غير المسماة تي سيارات الدفع الرباعي، يتم تقليل كثافة سطح fluorophore في الغشاء الآن جنبا إلى جنب، والزيادة الناتجة في دبي الوطني-PE مضان تقارير مدى الدهون خلط 10. كما فحص الجزء الأكبر من السهل إعداد وتحليل، وقد تم استخدامه على نطاق واسع لدراسة آليات بوساطة كمين الانصهار 10-14. ومع ذلك، لديها العديد من القيود، مثل انخفاض حساسية وضعف قرار الوقت. الأهم من ذلك، باعتبارها قياس الفرقة، فإنه المتوسطات النتائج على كل الأحداث مما يجعل التمييز بين الالتحام والانصهار، وكذلك الكشف عن وسيطة hemifusion الصعبة.
وعلى مدى عدة مجموعات العقد الماضي، بما في ذلك بلدنا، وضعت فحوصات جديدة لرصد الأحداث الانصهار في مستوى حويصلة واحدة 15-27. تستخدم ها وزملاؤه الخامس سيارات الدفع الرباعي المربوطة على سطح ومراقبة الانصهار مع حرية تي سيارات الدفع الرباعي 18،19. تم رصد الدهون خلط باستخدام الحنق بين زوج من fluorophores محددة الدهون مسريرا في الخامس وتي سيارات الدفع الرباعي، على التوالي، وذلك باستخدام الانعكاس الكلي الداخلي مضان (TIRF) المجهري 18. وفي وقت لاحق، استخدم مختبر Brunger من نوع دهون، تسمية واحدة جنبا إلى جنب مع علامة محتويات للكشف في وقت واحد من الدهون ومحتويات خلط 20،28. وشملت كلا من الدهون وعلامات محتويات في ارتفاع، تركيزات التبريد الذاتي. أدى اندماج مع سيارات الدفع الرباعي غير المسماة في مضان dequenching 20،28.
وتنصهر الآخرين الخامس سيارات الدفع الرباعي إلى طبقات ثنائية مستو تشكيلها مع تي الافخاخ 15-17،21-27،29. الهندسة المستوية من الهدف (تي فخ الحاوي) يحاكي أفضل طبقة ثنائية عملية الانصهار الفسيولوجية، الحويصلات المنحنية للغاية صغيرة مع غشاء البلازما المسطحة. مجموعة ستاينم المستخدمة الأغشية التي تغطي المسام تشكيلها مع تي الافخاخ، علقت اكثر من مسامية الركيزة نيتريد السيليكون والكشف عن الانصهار مع الفردية-سيارات الدفع الرباعي ضد استخدام ليزر متحد البؤر المسح المجهري 23. الآخرين وتستخدم الخامس سيارات الدفع الرباعي إلى طبقات ثنائية مستو تشكيلها مع تي الافخاخ، معتمد على ركيزة الزجاج 15-17،21،22،24-27،29. ميزة كبيرة لاستخدام طبقات ثنائية المدعومة (SBLs) هي أن TIRF المجهري يمكن استخدامها للكشف عن الالتحام والانصهار الأحداث مع ممتازة نسبة الإشارة إلى الضوضاء ودون تدخل من الخامس سيارات الدفع الرباعي الحرة، على الرغم من استخدام على microfluidics يوفر الدقة حدث واحد باستخدام معيار بعيدة الميدان epifluorescence المجهري 24.
ومصدر القلق الرئيسي هو ما إذا كان وكيف التفاعلات الركيزة طبقة ثنائية تؤثر دعم جودة طبقة ثنائية وعملية الاندماج. تقدم العمل في وقت مبكر استخدام SBLs مستو التي تم دعمها مباشرة على الزجاج أو الكوارتز الركيزة 15-17. وقدمت هذه SBLs عن طريق الامتزاز، انفجار، ونشر والانصهار الأغشية تي سيارات الدفع الرباعي على الركيزة. وقد أدرك ذلك قريبا، مع ذلك، أن حذف عنصر تي كمين رئيسي، SNAP25، من SBLs أعد بهذه الطريقة أدى إلى الخامس SUV الالتحام والانصهار حركية indistinguishable من تلك التي تم الحصول عليها باستخدام كامل تي الافخاخ 17. لأنه مطلوب SNAP25 تماما عن الانصهار في الجسم الحي 30،31، وضعت أهمية فسيولوجية من هذه المحاولات المبكرة موضع تساؤل. تغلبت مجموعة تام لهذا التحدي باستخدام تشكيل طبقة ثنائية الخاضعة لسيطرة دعم أفضل 21. وهي تستخدم انجميور-بلودجيت ترسب على النشرة الأولى خالية من البروتين من سيمبول تكنولوجيز، تليها انصهار أن أحادي الطبقة مع تي سيارات الدفع الرباعي 21. وأدى ذلك إلى الانصهار تعتمد-SNAP25.
لتجنب القطع الأثرية المحتملة المرتبطة طبقة ثنائية المدعومة مباشرة على ركيزة الزجاج دون الحاجة إلى استخدام أساليب انجميور-بلودجيت، Karatekin وآخرون قدم، بولي رطب (جلايكول الإثيلين) (PEG) وسادة لينة بين طبقة ثنائية والركيزة 24. أدى هذا التعديل أيضا في تعتمد-SNAP25 الانصهار 24. قد طبقات ثنائية خففت على طبقة البوليمر لينة من المعروف للحفاظ على الغشاء أفضلاستخدمت التنقل البروتين وظيفة 32، والدراسات الانصهار مع الفيروسات 33. وبالإضافة إلى ذلك، يبدو أن طبقات ثنائية مضاد للفيروسات للاحتفاظ بعض القدرة في تقرير المصير، شفاء وهي قوية جدا 34،35. أولا، واشتملت على جزء من المتاحة تجاريا سلاسل PEG، مرتبطة الدهون في غشاء تي سيارات الدفع الرباعي. عندما انفجرت هذه تي سيارات الدفع الرباعي وتشكل طبقة ثنائية مستو على ركيزة الزجاج وفرشاة PEG تغطي كلا منشورات من طبقة ثنائية مستو. لأنه هو الدافع وراء تشكيل مستو طبقة ثنائية الإذعان لسلاسل PEG المحيطة تي سيارات الدفع الرباعي على سطح الزجاج ماء، انفجار الحويصلية وتشكيل طبقة ثنائية مستو لا تعير اهتماما نسبيا لتكوين الدهون المستخدمة. ومع ذلك، عندما يتم إدراج كميات كبيرة من الكوليسترول في الدم، وزيادة خصائص متماسكة من سيارات الدفع الرباعي، وسيارات الدفع الرباعي قد لا تنفجر من تلقاء أنفسهم. إذا كان هذا هو الحال، ناضح صدمة أو ثنائي التكافؤ أيونات يمكن استخدامها للمساعدة على مستو تشكيل طبقة ثنائية 25.
وكما ذكر أعلاه، في هذه ا ف باملنهجية فرشاة PEG تغطي جانبي مستو، بدعم طبقة ثنائية. الفرشاة التي تواجه قناة تدفق الموائع الدقيقة يساعد على منع التصاق غير محددة من واردة الخامس سيارات الدفع الرباعي التي هي أيضا تغطيها عادة مع طبقة الربط. تشكيل المجمعات الخامس وتي كمين يبدأ من N-ترميني غشاء القاصي والعائدات في مراحل نحو المجالات غشاء الداني 36. ل-سيارات الدفع الرباعي الخامس للتفاعل مع تي سيمبول تكنولوجيز، والخامس والحاجة تي فخ N-تيرميني لتبرز فوق فرش PEG، والذي يبدو أن هذا هو الحال في ظل ظروف الفحص. يمكن تكييفها ارتفاع فرشاة لدراسة البروتينات الأخرى من الافخاخ من خلال تغيير كثافة من الدهون مضاد للفيروسات وPEG طول سلسلة 37،38. فائدة أخرى من فرش PEG تغطي الأسطح القريبة من طبقات ثنائية الصمامات هي أنها تحاكي بيئة مزدحمة من الأغشية الحيوية التي تكتظ 30،000-40،000 بروتينات الغشاء لا يتجزأ لكل متر مربع ميكرون 39. تماما مثل سلاسل PEG في هذا الاختبار، وREPUطبقة البروتين lsive تغطي الأغشية البيولوجية يجب أن تكون دفعت جانبا للسماح للاتصال بين اثنين من طبقات ثنائية فوسفورية للانصهار تحدث.
تستخدم قنوات تدفق الموائع الدقيقة في هذا الاختبار، كما أنها توفر مزايا فريدة من نوعها. أولا، تدفق ميكروفلويديك يتيح المزيد من ترسب موحد للتي سيارات الدفع الرباعي لنشر والصمامات لتشكيل تي SBL. الثانية، وحجم قناة صغيرة (<1 ميكرولتر) يقلل من استهلاك عينة. ثالثا، كميات صغيرة المطلوبة تسمح التجربة بأكملها التي ستجرى تحت تدفق مستمر. تدفق يزيل ضعيفة، ويفترض غير تحديدا، الالتزام الخامس سيارات الدفع الرباعي من SBL 16. كما يقيم كثافة ثابتة من الخامس سيارات الدفع الرباعي فوق تي سيمبول تكنولوجيز، وتبسيط التحليل الحركي 17. وأخيرا، وتتميز الحويصلات رست بسهولة من تلك الخطوط التي يحملها تدفق 25. الرابعة، وعدة قنوات ميكروفلويديك يمكن استخدامها على نفس ساترة، كل يحققون في حالة مختلفة. وهذا يسمح مقارنة الظروف الدورينانوغرام نفس العملية التجريبية. وقد استخدم نهج مماثل من قبل مجموعة فان Oijen لدراسة الاندماج بين فيروس الإنفلونزا، وSBLs خففت 33.
في TIRF المجهري، واضمحلال الأسي الحقل زائل (مع التحلل ثابت ~ 100 نانومتر) حدود مضان الإثارة إلى تلك الجزيئات التي هي في مكان قريب جدا من واجهة زجاجية عازلة. هذا يقلل من مساهمة جزيئات الفلورسنت التي هي أبعد، ويزيد من نسبة الإشارة إلى الضوضاء، ويتيح حساسية جزيء واحدة مع أوقات التعرض إطار 10-40 مللي ثانية. يؤدي مجال زائل أيضا إلى زيادة إشارة على الانصهار: كما نقل الدهون وصفت من سيارات الدفع الرباعي في سيمبول تكنولوجيز، وجدوا أنفسهم، في المتوسط، في حقل الإثارة أقوى. هذه الزيادة في مضان أقوى لالجسيمات الشحمية أكبر.
إذا تم استخدام الضوء المستقطب لتوليد مجال زائل، تساهم آثار إضافية للتغيرات في مضان على رransfer من التسميات من سيارات الدفع الرباعي في سيمبول تكنولوجيز. بعض الأصباغ الدهون لديها ثنائي القطب الانتقال الموجهة بزاوية متوسط يفضل فيما يتعلق طبقة ثنائية التي هي جزء لا يتجزأ منها. وهذا يخلق فرقا في مقدار مضان المنبعثة من fluorophores عندما يكونون في سيارات الدفع الرباعي مقابل SBL، لأن شعاع الاستقطاب سوف تثير الأصباغ في الأغشية اثنين بشكل مختلف. عن السابق، فإن شعاع الإثارة التفاعل مع ثنائيات الاقطاب الانتقال الموجهة نحو سيارات الدفع الرباعي كروية، في حين أن لهذا الأخير، وسوف تقتصر توجهات ثنائي القطب من قبل هندسة SBL مسطحة. على سبيل المثال، عند استخدام الصورة مستقطب الضوء الساقط (الاستقطاب وضعها الطبيعي لطائرة من الإصابة)، الإثارة أكثر كفاءة عندما الصبغة هي في SBL من في سيارات الدفع الرباعي لمواز الموجهة الدهون صبغ انتقال ثنائي القطب لغشاء 29،40 (مثل تلك التي الجاذبة أو لم 41-43). يظهر SUV مخدر مع مثل هذه fluorophore خافت عندما الاحواض على سيمبول تكنولوجيز (الشكل 7، Representat إيف نتائج). كما يفتح المسام الانصهار ويربط الأغشية سيارات الدفع الرباعي وSBL، تحقيقات الفلورسنت منتشر في SBL وتصبح أكثر عرضة للولع مجال زائل الاستقطاب يالي 25،27،29. وبالتالي، فإن إشارة مضان متكاملة حول الموقع الانصهار يزيد بشكل حاد خلال نقل الصبغة من سيارات الدفع الرباعي في SBL 27 (الشكل 3) والشكل (7). عاملا إضافيا يسهم في إشارة التغييرات التي ترافق الاندماج وdequenching من العلامات الفلورية كما هي المخفف أنها عندما نقل إلى SBL. مساهمة dequenching عادة طفيفة مقارنة آثار تسوس الميدانية والاستقطاب زائل في مقايسة الموصوفة هنا، لأنه فقط جزء صغير ( 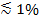 ) من الدهون وصفت.
) من الدهون وصفت.
الزيادة إشارة على الانصهار يمكن استغلالها لاستخلاص الخصائص الانصهار المسام بمقارنة ذلك الوقت،1 "SRC =" / ملفات / ftp_upload / 54349 / 54349eq2.jpg "/>، المطلوبة للدهون للهروب من خلال المسام التي هي قابلة للاختراق بحرية إلى الدهون إلى الافراج عن الوقت الفعلي،  . إذا جداول مرتين قابلة للمقارنة، ستكون النتيجة هي أن المسام يعرض مقاومة تذكر لتدفق الدهون. ومع ذلك، إذا كان الافراج عن الوقت الفعلي هو أطول بكثير من الوقت لاطلاق سراح محدودة الانتشار، وهذا من شأنه أن يشير إلى عملية، مثل الخفقان المسام، وتأخير الافراج عن الدهون. الافراج عن الوقت محدودة الانتشار،
. إذا جداول مرتين قابلة للمقارنة، ستكون النتيجة هي أن المسام يعرض مقاومة تذكر لتدفق الدهون. ومع ذلك، إذا كان الافراج عن الوقت الفعلي هو أطول بكثير من الوقت لاطلاق سراح محدودة الانتشار، وهذا من شأنه أن يشير إلى عملية، مثل الخفقان المسام، وتأخير الافراج عن الدهون. الافراج عن الوقت محدودة الانتشار، 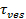 ، ويعتمد على حجم الجسيمات الشحمية الصمامات ودهون، الانتشارية. يتطلب تقديراتها هذه المعلمتين ليكون كميا. حساسية جزيء واحد من الفحص تسمح الانتشارية الدهون المراد قياسها من خلال تتبع عدة fluorophores الدهون واحد بعد إطلاق سراحهم في SBL لكل حدث الانصهار 26. حجم كل حويصلة الصماماتويمكن تقدير 27 عن طريق الجمع بين (ط) شدة صبغة الدهون واحد، (ب) التغير في مضان الكلي حول موقع رسو السفن بعد أن يتم نقل جميع fluorophores في سيمبول تكنولوجيز على الانصهار، (ج) المعروفة كثافة العلامات على سيارات الدفع الرباعي الدهون، و (د) من منطقة في الدهون. بالنسبة لكثير من الأحداث الانصهار، تم العثور على الأوقات الافراج عن الدهون الفعلية لتكون أبطأ بكثير مما كان متوقعا من قبل تحت سيطرة نشر الافراج عن 27، كما لوحظ سابقا على افتراض حجم SUV موحد 44. على افتراض التخلف الإفراج الدهون ومن المقرر أن المسام الخفقان، وهذا نموذج كمي يسمح تقدير "انفتاح المسام"، جزء من الوقت تبقى المسام مفتوحة أثناء الانصهار 27.
، ويعتمد على حجم الجسيمات الشحمية الصمامات ودهون، الانتشارية. يتطلب تقديراتها هذه المعلمتين ليكون كميا. حساسية جزيء واحد من الفحص تسمح الانتشارية الدهون المراد قياسها من خلال تتبع عدة fluorophores الدهون واحد بعد إطلاق سراحهم في SBL لكل حدث الانصهار 26. حجم كل حويصلة الصماماتويمكن تقدير 27 عن طريق الجمع بين (ط) شدة صبغة الدهون واحد، (ب) التغير في مضان الكلي حول موقع رسو السفن بعد أن يتم نقل جميع fluorophores في سيمبول تكنولوجيز على الانصهار، (ج) المعروفة كثافة العلامات على سيارات الدفع الرباعي الدهون، و (د) من منطقة في الدهون. بالنسبة لكثير من الأحداث الانصهار، تم العثور على الأوقات الافراج عن الدهون الفعلية لتكون أبطأ بكثير مما كان متوقعا من قبل تحت سيطرة نشر الافراج عن 27، كما لوحظ سابقا على افتراض حجم SUV موحد 44. على افتراض التخلف الإفراج الدهون ومن المقرر أن المسام الخفقان، وهذا نموذج كمي يسمح تقدير "انفتاح المسام"، جزء من الوقت تبقى المسام مفتوحة أثناء الانصهار 27.
كلما كان ذلك عمليا، فمن المهم لاختبار آليات الاندماج باستخدام كل من الدهون ومحتويات بطاقات القابلة للذوبان. على سبيل المثال، يمكن أن يؤخر الإفراج الدهون من خلال عمليات أخرى من الخفقان المسام، مثل تقييد انتشار الدهون من البروتينات الفخ الذي تحيط رانه المسام. إذا كانت هذه هي الحالة، ثم الإفراج عن محتويات سيسبق الإفراج عن تسميات الدهون، شريطة المسام كبيرة بما فيه الكفاية للسماح بمرور تحقيقات القابلة للذوبان. خلل أكثر أساسيا في نهج يمكن أن يكون في افتراض أن نقل الدهون المسمى إلى SBL يحدث من خلال المسام الانصهار الضيق الذي يربط بين SBL إلى الحويصلة التي احتفظت إلى حد كبير شكل قبل الانصهار. يمكن نقل الدهون في SBL ايضا ان ينتج من اتساع السريع للمسام الانصهار مع ما يصاحب ذلك، غاية الانهيار السريع من سيارات الدفع الرباعي في غشاء سيمبول تكنولوجيز، كما سبق واقترح بناء على بيانات الافراج عن الدهون وحدها 29. رصد كل الدهون ومحتويات الافراج في وقت واحد، فقد وجد أن العديد من المسام أعادت بعد إطلاق سراح جميع العلامات الدهون، ولكن احتفظت ببعض البضائع القابلة للذوبان من 27. هذا يدل على أن بعض الجسيمات الشحمية لا ينهار على الأقل في SBL بعد الانصهار، والتي نقل الدهون الصبغة في SBL يحدث من خلال المسام الانصهار. وبالإضافة إلى ذلك، لحدث IPID ومحتويات الافراج في وقت واحد 27، مما يجعل من غير المرجح أن تخلف الإفراج الدهون يرجع إلى إعاقة انتشار الدهون من البروتينات كمين المحيطة المسام 45.
بروتوكول الانصهار سيارات الدفع الرباعي SBL التي لم ترصد إطلاق محتويات قابلة للذوبان ونشرت من قبل Karatekin وروثمان 25. هنا، يتم تضمينها المزيد من التطورات الأخيرة، ورصد أي وقت واحد من الدهون والافراج عن محتويات وتقدير خصائص سيارات الدفع الرباعي، الدهون، والانصهار المسام 27. يبدأ البروتوكول مع تعليمات لإعداد الخلايا الموائع الدقيقة، التي تربط بولي (ميثيل siloxane) (PDMS) المطاط الصناعي كتلة تحتوي على أخاديد مع ساترة زجاج 25. بعد ذلك، يتم شرح إعداد الخامس سيارات الدفع الرباعي مع كل من الدهون ومحتويات علامات. القسمين 4 و 5 توفر تعليمات لتجميع الخلايا الموائع الدقيقة، وتشكيل SBLs في الموقع والتحقق من وجود عيوب وسيولة، وإدخال VSأوفس في تدفق الخلايا والكشف عن الأحداث الانصهار. ويقدم القسم 6 تعليمات لتحليل البيانات.
