膜融合为脂质和蛋白质,分泌,施肥,发展的细胞内运输所需的通用生物过程,和包膜病毒进入宿主生物体1-3。对于大多数细胞内聚变反应包括通过胞吐的激素和神经递质的释放,融合2脂质双层的能量由形成同源可溶性N-乙基马来酰亚胺-敏感因子附着蛋白受体 (SNARE)的蛋白质之间的四螺旋束的提供,锚定在囊泡(v-SNARE)和分别的靶膜(叔SNARE)4,。突触小泡胞吐作用是最严格调控的聚变反应和动作电位1,4,5到货后一毫秒内发生。的融合孔,两个熔断隔间之间的初始连接,可以闪烁打开和再密封或不可逆5-7扩大之前关闭多次。前者结果在短暂的“亲吻与运行”的融合,而后者导致完全融合。理事融合这两种模式和调节孔径闪烁机制之间的平衡因素不能很好地理解5,8。
SNARE蛋白所需的胞吐作用;突触囊泡融合是由神经毒素9时SNARE蛋白裂解取消。使用小单层囊泡(SUV的)堆积聚变实验表明,斯耐尔不仅必需的,但也足以驱动膜融合10。在此批量测定中,重构用v-SNARE蛋白(V-SUV)的SUV被掺杂有荧光磷脂(N – (7-硝基2-1,3-苯并恶二唑-4-基)-phosphoethanolamine(NBD-PE)和( N – (丽丝胺罗丹明B磺酰基)-phosphoethanolamine(LR-PE)和含叔SNARE蛋白(T-SUV)最初的v型的SUV NBD-PE的荧光被荧光共振能量转移淬无标签的小泡混合(FRET ),以LR-PE作为实验室ELED v型越野车与未标记叔SUV的融合,在现在结合膜的荧光面密度降低,在NBD-PE的荧光所产生的增加报告脂质混合10的程度。象散法是容易设置和分析,它已被广泛用来研究SNARE介导的融合10-14的机制。然而,它有一些限制,如低灵敏度和时间分辨率差。最重要的是,作为一个集合进行测定,平均结果在使半融合中间体难以检测对接和融合区分,以及所有事件。
在过去的十年几组,包括我们,已经开发了新的分析方法来监控在单级囊泡融合15-27事件。厦和他的同事用拴在表面上的V-SUV和监控他们与免费的T SUV的融合18,19。脂质混合是一对脂相连的荧光团之间的FRET使用EM监视层状分别在v型和叔越野车,使用全内反射荧光(TIRF)显微镜18。后来,Brunger的实验室使用单一脂质标签的物种连同内容标记物的脂质和内容混合20,28同时检测。既脂质和内容标记被包括在高,自猝灭浓度;融合SUV的未标记导致荧光脱猝灭20,28。
其他人已经融合v型越野车与T-SNARE蛋白15-17,21-27,29重组平面双层。目标(含T-SNARE)双层更好地模拟与一台质膜小,高度弯曲的囊泡的生理融合过程中的平面形状。该斯泰纳姆组采用与T-SNARE蛋白重组孔跨越膜,悬浮在多孔氮化硅基板,并检测使用激光共聚焦显微镜23个人的V型SUV的融合。其他˚F使用V型越野车与T-SNARE蛋白重组平面双层,支持在玻璃基板上15-17,21,22,24-27,29。使用支持的双层(SBLs)的巨大优点在于,TIRF显微镜可以用于检测具有优良信噪比并且在不脱离自由V形的SUV干扰对接和融合事件,虽然使用微流体使用还提供了单事件分辨率标准的远场荧光显微镜24。
一个主要问题是底物的相互作用双层是否以及如何影响支持双层质量和融合的过程。早期工作取得使用该直接支持的玻璃或石英衬底上15-17平面SBLs的。这些SBLs被吸附制成,爆裂,扩散和在衬底上叔SUV膜融合。它很快实现,然而,省略了密钥t-SNARE组分,SNAP25,从以这种方式制备SBLs导致V形的SUV对接和融合动力学indistinguishable使用完整的T-17 SNARE蛋白获得的。因为SNAP25 在体内 30,31绝对必需的融合,这些早期的尝试生理相关投入的问题。塔姆的研究小组通过使用更好的控制支持双层形成21战胜了这一挑战。它使用朗缪尔-布洛杰特沉积为SBL的无蛋白质的第一小叶,接着用叔-越野车21,其单层融合。这导致SNAP25依赖性融合。
以避免与直接支撑在玻璃基板上,而不需要使用朗缪尔-布洛杰特法的双层相关的潜在假象,Karatekin 等人介绍了柔软,水合聚(乙二醇)(PEG)的双层和基板24之间的缓冲。该变形也导致SNAP25依赖性融合24。垫着柔软的聚合物层上双层一直被称为更好地保存跨膜蛋白质的流动性和功能32,并与病毒融合33研究中使用。此外,聚乙二醇化双层似乎保留一定的能力进行自我修复,而且非常坚固34,35。首先,商购,脂质连接PEG链的一小部分被包括在叔SUV膜。当这些叔SUV的突发,并形成在玻璃基板上的平面双层,一个PEG刷覆盖平面双层的两个小叶。因为平面双层形成通过叔SUV的周围到所述亲水玻璃表面的PEG链的粘附驱动,脂质体破裂和平面双层形成相对不敏感,以所使用的脂质组合物。然而,当包括大量的胆固醇的,增加了越野车的粘结性时,越野车可以不自发破裂。如果是这种情况,可以采用渗透休克或二价离子,以帮助平面双层形成25。
如在本AP如上所述,proach的PEG刷覆盖所述平面的两侧,支持双层。面对微流体流动通道的刷有助于防止其也通常覆盖有聚乙二醇层传入v型越野车的非特异性粘附。 v型和叔SNARE复合体的形成从膜远侧N-末端并且前进朝向膜近端域36阶段开始。为V形的SUV与叔SBL中,v型和叔SNARE N-末端需要突出的PEG刷,这似乎是在测定条件下,上述情况下进行交互。刷高度可适于通过改变PEG化脂质的密度和PEG链长度37,38学习比斯耐尔其它蛋白质。所述PEG刷子覆盖定影双层的近侧表面中的另一个好处是,它们模仿它们填充有每平方微米39 30,000-40,000整合膜蛋白生物膜的拥挤的环境。就像在此测定的PEG链,所述REPU覆盖生物膜lsive蛋白层需要被推开以允许两个磷脂双层之间的接触为发生融合。
微流体流动通道在该测定中使用的,因为它们提供独特的优点。首先,微流体流使T-的SUV更均匀的沉积扩散和融合形成了T-SBL。其次,小声道音量(<1微升)减少样品消耗。第三,所需的小体积允许下恒定流量进行的整个实验。流动弱除去,大概非特异性,从SBL 16附着v型越野车。它还维护V型的SUV的T-SBL以上,简化动力学分析17的密度不变。最后,停靠囊泡容易从由流25进行自由的区分。第四,多个微流体通道可以在相同的盖玻片一起使用,每个探测不同的条件。这使得杜里条件比较纳克相同的实验运行。类似的方法已被用于面包车Oijen组研究流感病毒和缓冲SBLs 33之间的融合。
在TIRF显微镜,渐逝场的指数衰减(以衰减常数〜100nm)的界限荧光激发到那些在玻璃 – 缓冲界面的非常接近的分子。这最小化是更远离荧光分子的贡献,增加信噪比,并且允许用10-40毫秒的帧的曝光时间单分子灵敏度。渐逝场也导致在融合的信号增加:从SUV标记的脂质转移到SBL,他们发现自己,平均而言,在更强的激励场。这增加了荧光较大的脂质体更强。
如果偏振光是用来产生渐逝场,附加效果有助于对T中的荧光变化从SUV到SBL标签的转让(BOT)。某些脂质染料具有与相对于它们所嵌入的双层的优选平均角度定向的跃迁偶极。这将在通过荧光团时,他们在SUV 兑 SBL发射的荧光的量的差异,由于偏振光分束将在两个膜激发染料不同。对于前者,激发光束将与周围的球形的SUV取向过渡偶极子相互作用,而对于后者,偶极取向将由扁平SBL几何局限。例如,当S偏振入射的光(偏振光垂直于入射平面)时,激发是更有效率的,当染料在SBL比在SUV脂质染料跃迁偶极方向平行于膜29,40一(如直接投资收益还是没41-43)。掺杂了这样的荧光团SUV似乎暗淡时,码头上SBL( 图7,Representat香港专业教育学院结果)。作为融合孔打开,并连接在SUV和SBL膜,荧光探针扩散到SBL和变得更容易被s偏振光渐逝场25,27,29激发。因此,周围的融合位点整合的荧光信号从所述的SUV染料转移到SBL 27( 图3和图7)期间急剧增加。有助于信号伴随融合脱猝灭的荧光标记的时转移到SBL因为它们稀释变化的另外一个因素。相比,在这里描述的测定消逝场衰减和极化效应脱猝灭的贡献通常是次要的,因为只有一小部分( 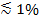 )脂质的标记。
)脂质的标记。
在融合信号增加可以利用通过比较时间来推断融合孔性质,1“SRC =”/文件/ ftp_upload / 54349 / 54349eq2.jpg“/>,对于脂质通过的孔是自由可渗透脂质实际释放时间逃逸要求 。如果两个时间尺度是可比,将它的结论是,孔呈现给脂质流动阻力小。然而,如果实际的释放时间是显著比时间为扩散限制释放更长的时间,这将表明一个过程中,如孔隙抖动,延迟脂质释放。扩散限制释放时间,
。如果两个时间尺度是可比,将它的结论是,孔呈现给脂质流动阻力小。然而,如果实际的释放时间是显著比时间为扩散限制释放更长的时间,这将表明一个过程中,如孔隙抖动,延迟脂质释放。扩散限制释放时间, 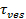 取决于定影脂质体和脂质扩散的大小;其估计需要这两个参数来量化。测定的单分子灵敏度允许脂质扩散到其释放到SBL为每个聚变事件26之后追踪几个单脂的荧光团测定。每一熔断囊泡的大小可通过将(i)的单脂质染料的强度,(ii)在所有荧光团在融合转移到SBL中围绕一个停靠站点的总荧光变化,(iii)该已知标记的SUV的密度来估计27脂质,和(iv)每脂质的区域。对于许多融合事件中,发现了实际脂质释放时间要慢得多比由扩散控制的释放27预期,如先前指出的假定均匀的SUV大小44。假设脂质释放迟缓是由于毛孔闪烁,定量模型允许“毛孔开放”的估计,时间的部分孔隙依然融合27期间开放。
取决于定影脂质体和脂质扩散的大小;其估计需要这两个参数来量化。测定的单分子灵敏度允许脂质扩散到其释放到SBL为每个聚变事件26之后追踪几个单脂的荧光团测定。每一熔断囊泡的大小可通过将(i)的单脂质染料的强度,(ii)在所有荧光团在融合转移到SBL中围绕一个停靠站点的总荧光变化,(iii)该已知标记的SUV的密度来估计27脂质,和(iv)每脂质的区域。对于许多融合事件中,发现了实际脂质释放时间要慢得多比由扩散控制的释放27预期,如先前指出的假定均匀的SUV大小44。假设脂质释放迟缓是由于毛孔闪烁,定量模型允许“毛孔开放”的估计,时间的部分孔隙依然融合27期间开放。
每当实际,以测试同时使用脂质和可溶性内容标签融合的机制是很重要的。例如,脂质释放可以通过比孔隙闪烁等工艺,如由SNARE蛋白是环绕吨脂质扩散的限制被延迟他孔隙。如果是这样的话,然后松开的内容将先于脂质标签释放,所提供的孔径是大到足以允许可溶性探针通道。在方法的一个更基本的缺陷可能是通过一个狭窄的融合孔的SBL连接到囊泡已经基本上保留了其融合前形状发生标记的脂质转移到SBL的假设。脂质转移到SBL也可以从融合孔与随之而来的,超极本的SUV进入SBL膜的迅速崩溃的快速散瞳造成的,正如上面所提到仅仅基于29脂质发布的数据。同时监测血脂和内容同时发布,发现许多气孔释放他们所有的脂质标签后重新密封,但保留了他们的一些可溶性的货物27。这表明,至少一些脂质体不塌陷到SBL融合之后,并且该脂质染料转移到SBL通过融合孔发生。此外,LIPID和内容,同时发布27发生,使得它不可能脂质释放迟缓,是由于受到周围孔45 SNARE蛋白阻碍脂质扩散的。
未监测溶于内容发布一款SUV-SBL融合协议之前由Karatekin和罗斯曼25公布。在这里,最近的事态发展都包括在内,脂质即同步监测和内容发布和SUV,脂质,并融合孔特性27的推定。该协议开始于制备微流体细胞,通过结合的聚(二甲基硅氧烷)制成(PDMS)弹性体包含块槽用玻璃盖玻片25的指令。接着,制备V-SUV的与两个脂质和内容标记进行说明。第4和5用于组装微流体细胞, 原位形成的SBLs并检查缺陷和流动性,引入VS的提供指令UVS到流动细胞和融合事件的检测。部分6提供用于数据分析的指令。
