A fusão da membrana é um processo biológico universal necessário para o tráfico intracelular de lípidos e proteínas, secreção, fertilização, desenvolvimento, e envolveu a entrada do vírus em organismos hospedeiros 1-3. Para a maioria das reacções de fusão intracelular incluindo libertação de hormonas e de neurotransmissores via exocitose, a energia para fundir duas bicamadas lipídicas é proporcionado pela formação de um feixe de quatro hélices entre proteínas de receptor cognato solúvel N-etilmaleimida-sensível proteína factor de fixação (SNARE), ancorada na a vesícula (v-SNARE) e da membrana alvo (T-SNARE) 4, respectivamente. Synaptic exocitose da vesícula é a reação de fusão mais fortemente regulada e ocorre dentro de um milésimo de segundo depois da chegada de um 1,4,5 potencial de ação. O poro de fusão, a ligação inicial entre os dois compartimentos de fusão, pode piscar aberta e fechada várias vezes antes de selar ou expandir irreversivelmente 5-7. Os resultados anterioresem transitória, "Kiss & run" fusion, enquanto o segundo leva a fusão completa. Fatores que regem o equilíbrio entre estes dois modos de fusão e mecanismos que regulam cintilação poros não são bem compreendidos 5,8.
SNARE proteínas são necessárias para a exocitose; fusão das vesículas sinápticas é abolida por quebra de SNAREs por neurotoxinas 9. Experiências de fusão em massa usando pequenas vesículas unilamelares (SUVs) mostrou que SNAREs não só necessário, mas também suficiente para conduzir a fusão da membrana 10. Neste ensaio a granel, SUVs reconstituído com v-SNARE (V-SUV) foram dopados com fosfolípidos fluorescentes (N – (7-nitro-2-1,3-benzoxadiazol-4-il) -phosphoethanolamine (NBD-PE) e ( N -. (lisamina rodamina B sulfonil) -phosphoethanolamine (LR-PE) e misturado com vesículas não marcados contendo t-SNAREs (t-SUV) Inicialmente, a fluorescência do NBD-PE no v-SUVs é extinta por Förster transferência de energia de ressonância (FRET ) para LR-PE. Como laboratórioELED v-SUVs fundir-se com t-SUVs não marcados, a densidade da superfície fluoróforo na membrana agora combinada é reduzido e o aumento resultante na fluorescência de NBD-PE relata a extensão de lípido de mistura 10. Como o ensaio é fácil de grandes quantidades definir-se e analisar, que tem sido amplamente utilizada para estudar os mecanismos de fusão mediada por SNARE 10-14. No entanto, tem várias limitações, como a baixa sensibilidade e baixa resolução tempo. O mais importante, como um conjunto de medição, ela é em média resultados de todos os eventos que fazem a discriminação entre o encaixe e de fusão, bem como a detecção de intermediários hemifusão difíceis.
Durante a última década vários grupos, incluindo o nosso, desenvolveram novos ensaios para monitorar eventos de fusão no único nível vesícula 15-27. Ha e seus colegas usaram v-SUVs amarrados em uma superfície e monitorados sua fusão com livre t-SUVs 18,19. mistura lipídica foi monitorizada utilizando FRET entre um par de fluoróforos ligados a lípidos inalojado no V e t-SUVs, respectivamente, através de microscopia de fluorescência de reflexão total interna (TIRF) 18. Mais tarde, o laboratório de Brunger utilizada uma única espécie de rótulo de lípidos juntamente com um marcador de conteúdos para a detecção simultânea de lípido de mistura e os conteúdos 20,28. Tanto o lipídico e os marcadores de conteúdo foram incluídos em concentrações elevadas, auto-extingue; fusão com SUVs não marcados resultou em fluorescência dequenching 20,28.
Outros têm fundido v-SUVs para bicamadas planar reconstituídos com t-SNAREs 15-17,21-27,29. A geometria planar do alvo (T-SNARE contendo) bicamada imita melhor o processo de fusão fisiológica de pequenas vesículas, altamente curvadas com uma membrana de plasma plana. O grupo Steinem empregada membranas de abrangência dos poros reconstituídos com t-SNAREs, suspensas sobre um substrato de nitreto de silício poroso e detectado fusão com v-SUVs individuais usando microscopia confocal de varrimento laser 23. outros fusados v-SUVs para bicamadas planar reconstituídos com t-SNAREs, apoiada sobre um substrato de vidro 15-17,21,22,24-27,29. A grande vantagem do uso de camadas duplas suportados (SBLs) é que a microscopia TIRF pode ser usado para detectar atracação e de fusão eventos com excelente relação sinal-ruído e sem interferência de V-SUVs livres, embora usando microfluídica também fornece resolução single-evento usando microscopia de epifluorescência-campo distante padrão de 24.
Uma preocupação importante é se, e como interacções substrato-bicamada suportada afectar a qualidade da bicamada e o processo de fusão. Os primeiros trabalhos feito uso de SBLs planares que foram suportados directamente sobre um substrato de vidro ou de quartzo 15-17. Estes SBLs foram feitas por adsorção, a rebentar, espalhando e fusão de membranas t-SUV sobre o substrato. Foi logo percebeu, porém, que omitir um componente-chave t-SNARE, SNAP25, de SBLs preparados desta maneira resultou em v-SUV de atracação e de fusão cinética indistinguishable daqueles obtidos utilizando a completa t-SNAREs 17. Porque SNAP25 é absolutamente necessária para a fusão in vivo 30,31, a relevância fisiológica desses primeiras tentativas foi posta em causa. O grupo de Tamm superou este desafio usando a formação de bicamada melhor controlada suportado 21. Usou-se a deposição de Langmuir-Blodgett, pela primeira folheto livre de proteínas de SBL, seguido por fusão de que monocamada com t-SUVs 21. Isto resultou na fusão dependente de SNAP25.
Para evitar artefactos potenciais associados a uma bicamada suportada directamente sobre um substrato de vidro, sem necessidade de utilizar métodos de Langmuir-Blodgett, Karatekin et ai. Introduziu um poli hidratado (etileno glicol) de apoio macio, (PEG) entre a bicamada e o substrato 24. Esta modificação também resultou na fusão dependente da SNAP25 24. Bicamadas almofadadas em uma camada de polímero macio tinha sido conhecido para melhor preservar transmembranamobilidade e função das proteínas de 32, e tinha sido utilizado em estudos de fusão com vírus 33. Além disso, bicamadas PEGilados parecem manter alguma capacidade de se auto-curar e são muito robustos 34,35. Em primeiro lugar, uma fracção de cadeias de PEG disponíveis comercialmente, ligados em lípidos incluídos na membrana t-SUV. Quando estes t-SUVs rebentar e formar uma bicamada planar num substrato de vidro, uma escova de PEG abrange ambos os folhetos da bicamada planar. Como a formação de bicamada planar é impulsionado por intermédio de adesão das cadeias de PEG circundantes t-SUVs sobre a superfície de vidro hidrófilo, rebentamento de lipossomas e formação em bicamada planar são relativamente insensíveis à composição de lípido utilizado. No entanto, quando grandes quantidades de colesterol estão incluídos, aumentando as propriedades coesivas dos SUVs, as SUVs não pode rebentar espontaneamente. Se este for o caso, os iões divalentes ou choque osmótico pode ser empregado para ajudar a formação planar de duas camadas 25.
Como mencionado acima, nesta approach uma escova de PEG cobre ambos os lados do cartão planar, suportado bicamada. A escova de frente para o canal de fluxo de microfluidos ajuda a evitar a aderência não específica da v-SUVs de entrada que também são geralmente cobertos com uma camada de PEG. A formação de complexos e t-V- SNARE começa a partir do terminal N da membrana distal e procede em etapas para com os domínios membrana proximal 36. Para os v-SUVs para interagir com o t-SBL, a necessidade e v- t-SNARE terminais N a projectar-se acima das escovas de PEG, o que parece ser o caso nas condições do ensaio. Altura da escova pode ser adaptada para estudar outras proteínas que SNARE, variando a densidade de lipidos PEGilados e o comprimento da cadeia de PEG 37,38. Um outro benefício das escovas de PEG que cobre as superfícies proximais das bicamadas de fusão é que eles imitam o ambiente de aglomerado de membranas biológicas, que são embalados com 30.000-40.000 proteínas de membrana integrais por quadrado 39 micron. Assim como as cadeias de PEG Neste ensaio, o repucamada de proteína lsive abrangendo membranas biológicas tem de ser empurrado de lado para permitir o contacto entre as duas bicamadas de fosfolípidos para a fusão ocorresse.
canais de microfluidos de fluxo são utilizados neste ensaio, uma vez que oferecem vantagens únicas. Primeiro, o fluxo microfluídico permite a deposição uniforme mais de t-SUVs para espalhar e se fundem para formar o t-SBL. Em segundo lugar, o volume do canal de pequeno (<1 ul) minimiza o consumo da amostra. Em terceiro lugar, as pequenas quantidades necessárias de permitir que todo o experimento ser conduzida sob fluxo constante. Fluxo remove fracamente, presumivelmente não-especificamente, aderiram v-SUVs da SBL 16. Ele também mantém uma densidade constante de v-SUVs acima do t-SBL, simplificando a análise cinética 17. Finalmente, vesículas ancoradas são facilmente distinguidos dos mais livre transportados pelo fluxo 25. Em quarto lugar, vários canais de microfluidos pode ser utilizado na mesma lamela, cada sondagem de uma condição diferente. Isto permite a comparação das condições de during mesma corrida experimental. Uma abordagem semelhante foi usada pelo grupo van Oijen para estudar fusão entre vírus influenza e SBLs almofadadas 33.
Em microscopia TIRF, o decaimento exponencial do campo evanescente (com um decaimento constante de ~ 100 nm) limites de excitação de fluorescência para aquelas moléculas que são muito próximo da interface vidro-tampão. Isso minimiza a contribuição de moléculas fluorescentes que estão mais longe, aumenta a relação sinal-ruído, e permite sensibilidade única molécula com tempos de exposição de quadro de 10-40 ms. O campo evanescente também leva a um aumento de sinal em cima de fusão: como a transferência de lípidos marcados a partir do SUV para o SBL, encontram-se, em média, em um campo de excitação mais forte. Este aumento na fluorescência é mais forte para os lipossomas maiores.
Se a luz polarizada é usada para gerar o campo evanescente, efeitos adicionais contribuir para alterações na fluorescência após transferência de etiquetas do SUV na SBL. Alguns corantes de lípidos tem um dipolo de transição orientada com um ângulo médio preferido em relação à camada dupla em que eles são incorporados. Isto cria uma diferença de a quantidade de fluorescência emitida pelos fluoroforos quando estão no SUV versus a SBL, uma vez que o feixe polarizado vai excitar corantes nas duas membranas de forma diferente. Para o primeiro, o feixe de excitação vai interagir com dipolos orientados de transição em torno do SUV esférica, ao passo que para o segundo, orientações dipolo será confinado pela geometria plana SBL. Por exemplo, quando é usado s-luz polarizada incidente (polarizado normal ao plano de incidência), a excitação é mais eficiente quando o corante é no SBL do que no SUV por um corante de lípidos transição dipolo orientado paralelamente à membrana 29,40 (como a de DII ou fez 41-43). Um SUV dopado com tal fluoróforo aparece dim quando se dirige para o SBL (Figura 7, Representat ive Resultados). Como um poro de fusão abre e liga as membranas SUV e SBL, sondas fluorescentes difundem-se para a SBL e tornar-se mais susceptível de ser excitado pelo campo evanescente s-polarizados 25,27,29. Por conseguinte, o sinal de fluorescência integrada em torno do local de fusão aumenta acentuadamente durante a transferência de corantes a partir de SUV para a SBL 27 (Figura 3 e Figura 7). Um factor adicional que contribui para sinalizar mudanças que acompanham fusão é dequenching de etiquetas fluorescentes, como eles são diluídos quando transferido para a SBL. A contribuição de dequenching é geralmente menor em comparação com efeitos evanescentes decaem campo e polarização no ensaio descrito aqui, porque apenas uma pequena fração ( 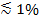 ) Dos lípidos são rotulados.
) Dos lípidos são rotulados.
O aumento do sinal mediante a fusão pode ser explorada para deduzir propriedades de poros de fusão através da comparação do tempo,1 "src =" / files / ftp_upload / 54349 / 54349eq2.jpg "/>, necessário para um lipídio de escapar através de um poro que é livremente permeável à lipídios com o tempo de lançamento real,  . Se as duas escalas de tempo são comparáveis, ao que se concluir que a poro apresenta pouca resistência ao fluxo de lípido. No entanto, se o tempo efectivo de libertação é significativamente mais longo do que o tempo para a libertação limitada por difusão, isso indicaria um processo, tal como tremulação de poro, retardador de libertação de lípidos. O tempo de liberação limitada por difusão,
. Se as duas escalas de tempo são comparáveis, ao que se concluir que a poro apresenta pouca resistência ao fluxo de lípido. No entanto, se o tempo efectivo de libertação é significativamente mais longo do que o tempo para a libertação limitada por difusão, isso indicaria um processo, tal como tremulação de poro, retardador de libertação de lípidos. O tempo de liberação limitada por difusão, 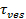 , Depende do tamanho do lipossoma de fusão e de lípidos-difusividade; sua estimativa exige estes dois parâmetros para ser quantificado. A sensibilidade única molécula do ensaio permite diffusivity lipídica a ser medido através do rastreamento vários fluoróforos única de lipídios após a sua libertação no SBL para cada evento de fusão 26. O tamanho de cada vesícula fusãopode ser cerca de 27 por combinação de (I) a intensidade de um único corante lípido, (ii) a alteração na fluorescência total em torno de um local de ancoragem depois de todos os fluoróforos são transferidos para a SBL mediante fusão, (iii) a densidade de rotulagem conhecidos de SUV lípidos, e (iv) a superfície por lípido. Para muitos eventos de fusão, os tempos de libertação de lípidos reais foram encontrados para ser muito mais lento que o esperado pela libertação de difusão controlada 27, como foi observado anteriormente assumindo SUV tamanho uniforme 44. Assumindo retardamento da liberação de lipídios é devido a pore cintilação, um modelo quantitativo permite a estimativa de "abertura dos poros", a fração de tempo do poro permanece aberta durante a fusão 27.
, Depende do tamanho do lipossoma de fusão e de lípidos-difusividade; sua estimativa exige estes dois parâmetros para ser quantificado. A sensibilidade única molécula do ensaio permite diffusivity lipídica a ser medido através do rastreamento vários fluoróforos única de lipídios após a sua libertação no SBL para cada evento de fusão 26. O tamanho de cada vesícula fusãopode ser cerca de 27 por combinação de (I) a intensidade de um único corante lípido, (ii) a alteração na fluorescência total em torno de um local de ancoragem depois de todos os fluoróforos são transferidos para a SBL mediante fusão, (iii) a densidade de rotulagem conhecidos de SUV lípidos, e (iv) a superfície por lípido. Para muitos eventos de fusão, os tempos de libertação de lípidos reais foram encontrados para ser muito mais lento que o esperado pela libertação de difusão controlada 27, como foi observado anteriormente assumindo SUV tamanho uniforme 44. Assumindo retardamento da liberação de lipídios é devido a pore cintilação, um modelo quantitativo permite a estimativa de "abertura dos poros", a fração de tempo do poro permanece aberta durante a fusão 27.
Sempre que possível, é importante para testar mecanismos de fusão usando tanto conteúdo etiquetas solúveis lipídico e. Por exemplo, a liberação de lipídios pode ser retardado por outros que cintilação poros processos, como a restrição da difusão de lípidos pelas proteínas SNARE que cercam tele poros. Se este fosse o caso, em seguida, solte do conteúdo precederia de suporte de etiquetas de lipídios, desde o poro é grande o suficiente para permitir a passagem de sondas solúveis. Uma falha mais fundamental na abordagem poderia ser sob a suposição de que a transferência de lípidos marcados para a SBL ocorre através de um poro de fusão estreita que liga a SBL a uma vesícula que manteve a sua forma, em grande parte de pré-fusão. Transferência de lípides para a SBL também pode resultar de uma rápida dilatação dos poros de fusão com uma concomitante, extremamente rápido colapso do SUV na membrana SBL, como previamente sugerido com base em dados de libertação de lípidos sozinho 29. Monitoramento tanto lipídica e conteúdo liberar simultaneamente, verificou-se que muitos poros fechados de novo depois de libertar todos os seus rótulos de lipídios, mas manteve alguma da sua carga solúvel 27. Isto indica que, pelo menos alguns lipossomas não colapsam para a SBL após a fusão, e que a transferência de corante de lípidos no SBL ocorre através de um poro de fusão. Além disso, lIPID e conteúdo de libertação ocorreu simultaneamente 27, tornando improvável que retardamento da libertação de lípidos era devido ao impedimento de difusão de lípidos pelas proteínas SNARE circundantes do poro 45.
Um protocolo de fusão SUV-SBL que não monitorar liberação conteúdos solúveis foi publicado anteriormente pelo Karatekin e Rothman 25. Aqui, os desenvolvimentos mais recentes são incluídos, monitoramento ou seja simultânea de lipídios e conteúdo liberar e estimativa de propriedades SUV, lipídios, e de poros de fusão 27. O protocolo começa com instruções para preparar as células de microfluidos, realizadas por ligação de um poli (dimetilsiloxano) (PDMS) elastómero de blocos contendo ranhuras com uma lamela de vidro de 25. Em seguida, preparação de v-SUVs com tanto lipídica e conteúdo marcadores é explicado. Secções 4 e 5 fornecem instruções para a montagem das células microfluídicos, formando os SBLs in situ e verificar se há defeitos e fluidez, introdução de vSUVS para as células de fluxo e detecção de eventos de fusão. Secção 6 fornece instruções para a análise de dados.
