Thymic slices support analysis of different aspects of T cell development such as positive and negative selection. For successful experiments, the quality of the thymic slice is paramount. Thus, thymic slices should be examined to ensure the integrity of the thymic tissue and that the agarose surrounding the thymic slice is intact (Figure 1A). Surface tension can be compromised when the agarose is damaged causing a significant decrease in the number of thymocytes that migrate into the tissue. Thus, thymic slices with indications of tissue damage or nicks/tears in the agarose should be discarded (Figure 1B).
TCR transgenic (tg) mice are often employed to study thymic selection due to the expression of a defined, functional TCR on the surface of every thymocyte that increases the frequency of positive selection over polyclonal populations. In order to capture each stage of positive selection, pre-selection TCR tg thymocytes can be overlaid atop thymic slices, and the development of mature, CD4+ or CD8+ SP T cells followed over time by flow cytometry40. Thymocytes isolated from MHC class I-restricted OT-I TCR tg Rag1-/- β2m-/- mice are arrested at the DP stage prior to thymic selection due to the necessity of β2m to associate with and stabilize the MHC class I heavy chain41,42. When overlaid onto β2m-/- thymic slices, the pre-selection OT-I TCR tg thymocytes remain at the DP stage and do not generate significant CD8+ SP T cells (Figure 2A, left). In contrast, when overlaid on WT thymic slices containing endogenous selecting ligands, the ability of this model to support positive selection is evidenced by CD8+ T cell development at 72 hr (Figure 2A, right). Quantification of CD8+ T cell development as a read-out of positive selection is shown in Figure 2B.
Thymic slices can also be used to study negative selection. A large proportion of negative selection occurs in response to ubiquitous antigen16. To model negative selection to ubiquitous antigen, total thymocytes from OT-I TCR tg Rag1-/- and wild-type (WT) mice were labelled with CFSE and CTV, respectively, then mixed in a 1:1 ratio and overlaid onto WT slices (Figure 3A). After washing off excess thymocytes, the thymic slices were placed in complete RPMI-1640 media with or without 1 nM of the cognate, agonist peptide of the OT-I TCR, SIINFEKL (OVA peptide). All MHC class I expressing cells will present the antigen, modeling the presentation of ubiquitously expressed antigen in the thymus. After 24 hr of incubation at 37 °C, the proportion of OT-I TCR tg cells (% live CFSE+) to WT control cells (% live CTV+) was compared by flow cytometry (Figure 3B). A decrease in the relative proportion of OT-I TCR tg cells represents deletion of OTI TCR tg cells in response to OVA peptide.
It is important to note that due to heterogeneity in the size of thymic slices as well as cell entry into thymic slices, one should include appropriate analysis and internal controls to normalize for these variables. Here, WT cells mixed with OT-I TCR tg cells and overlaid atop slices were used as an internal control as this population should not be significantly affected by the presence or absence of OVA peptide. The extent of negative selection of OTI TCR tg cells was quantified based on proportions rather than absolute numbers. First, the ratio between OT-I TCR tg (% live CFSE+) to WT (% live CTV+) cells on each WT thymic slice in the presence or absence of OVA peptide was calculated. Each of these ratios was then divided by the mean of the ratio between OT-I TCR tg (% live CFSE+) to WT (% live CTV+) cells on WT slices in the absence of OVA peptide. On average, 80% of the OT-I TCR tg cells were depleted in response to ubiquitous antigen as depicted in Figure 3C.

Figure 1: Representative images of thymic slices prepared from a vertically embedded thymic lobe. (A) Good quality slice with the embedded tissue and surrounding agarose intact. (B) Poor quality slice with damage due to tearing of the embedded tissue during slicing. Please click here to view a larger version of this figure.
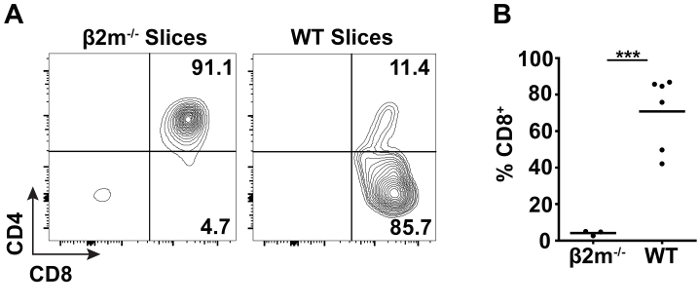
Figure 2: Flow cytometric analysis of positive selection in thymic slices. OT-I TCR tg Rag1-/- β2m-/- thymocytes were labelled with CFSE and overlaid onto non-selecting β2m-/- or selecting WT slices. After 72 hr of incubation at 37 °C, CD8+ T cell development was analyzed by flow cytometry. (A) Representative data of cell surface CD4 and CD8 expression on live CFSE+ TCRβhigh thymocytes from β2m-/- and WT slices. (B) Quantification of live CFSE+ TCRβhigh CD8+ T cell development on WT slices as compared to non-selecting controls. Each dot represents an individual thymic slice and the line represents the mean. (***p < 0.001, unpaired t test) Please click here to view a larger version of this figure.
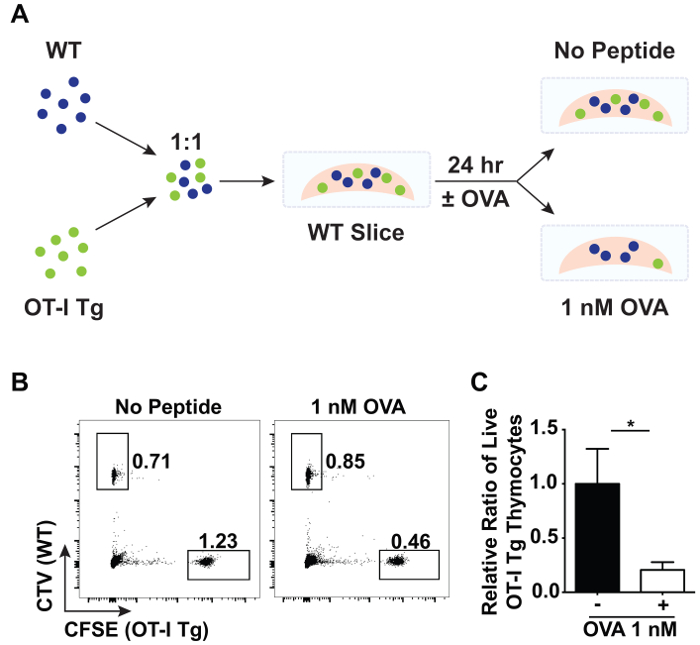
Figure 3: Flow cytometric analysis of negative selection in thymic slices. A 1:1 ratio of total CFSE-labeled OT-I TCR tg Rag1-/- and CTV-labeled WT thymocytes were overlaid on WT slices in the presence or absence of the OVA peptide, SIINFEKL. After 24 hr of incubation at 37 °C, the relative proportions of overlaid cells were analyzed by flow cytometry. (A) A schematic of the experimental set-up showing an overlay of CTV-labeled WT cells (blue) mixed 1:1 with CFSE-labeled OT-I TCR tg cells (green) onto a WT thymic slice in the presence or absence of OVA peptide. (B) Representative data depicting the proportion of live CFSE+ OT-I TCR tg cells and CTV+ WT cells from slices that were incubated with or without OVA peptide. (C) Data was normalized between slices, and the relative ratio of live CFSE+ OT-I TCR tg cells to live CTV+ WT cells is shown. Error bars indicate standard deviation. N = 3 for each condition. (*p < 0.05, unpaired t test). Please click here to view a larger version of this figure.
