وتقدم هذه الورقة فحص موثوق للإبلاغ عن التعبير سطح الخلية النسبي للبروتينات غشاء مثل القنوات الأيونية التي أعرب عنها في خلايا المؤتلف باستخدام تقنية التدفق الخلوي القائمة. القنوات الأيونية هي تشكيل المسام بروتينات غشاء التي هي المسؤولة عن السيطرة على الإشارات الكهربائية عن طريق النابضة تدفق الأيونات عبر غشاء الخلية. يتم تصنيفها من قبل آلية تفعيل، والطبيعة، والانتقائية الأنواع أيون تمر عبر المسام حيث يتم ترجمة ما. في مستويات الخلايا والأنسجة، وتدفقات أيون العيانية من خلال القنوات الأيونية هي نتاج الخصائص الفيزيائية الحيوية 1 (النابضة وتخلل)، والكيمياء الحيوية (الفسفرة)، ونشوء حيوي (التوليف، بالغليكوزيل، والاتجار، وتدهور). كل من هذه العمليات هي فريدة من نوعها لكل نوع من القنوات الأيونية وهو الأمثل لأداء الدور الفسيولوجي للقناة أيون. ونتيجة لذلك، وتعديلات في أي من هذه العمليات ضبطها غرامة من خلالالموروثة أو التعديل الوراثي، وغالبا ما يشار إليها باسم "اعتلال القنوات"، يمكن أن يؤثر سلبا على التوازن الخلية. ومن المهم التأكيد على أن توفير "الحق" كمية من القنوات الأيونية على سطح الخلية أمر بالغ الأهمية لتوازن الخلايا. حتى زيادات صغيرة (ربح من وظيفة) وانخفاض صغيرة (الخسارة من وظيفة) في نشاط القناة الايونية لديها القدرة على التسبب في أمراض خطيرة على مدى العمر. عيوب في تسليم سطح الخلية من القنوات الأيونية الناضجة هي من العوامل الهامة في العديد من channelopathies، مثل التليف الكيسي (القناة الايونية CFTR) (2) وعدم انتظام ضربات القلب من شكل طويل كيو تي متلازمة (قنوات البوتاسيوم في القلب) 3.
ترتبط Channelopathies مع الموت المفاجئ القلب 4. ويعتقد أن انتشار في جميع أنحاء العالم الحالي لجميع channelopathies القلب أن يكون على الأقل 1: 2،000-1: 3000 في الفردي 5، ومسؤولة عن حوالي نصف المفاجئ عدم اتساق نبضات القلب كاليفورنيا موت القلبإس إي إس 6. ومن المعروف أن الخلل في القلب الجهد بوابات sodium-، البوتاسيوم، والكالسيوم القنوات الأيونية الانتقائية للعب دورا رئيسيا في هذه العملية. مطلوب L من نوع كا V 1.2 قناة الكالسيوم الجهد بوابات لبدء متزامنة القلب تقلص العضلات. وقلبية L من نوع كا V 1.2 قناة هو بروتين معقد متعدد فرعية مؤلفة من تشكيل المسام الرئيسية الكالسيوم فرعية الخامس α1 والكالسيوم الخامس ß والكالسيوم الخامس α2δ1 مفارز المساعدة 7-12. لاحظ أنه مطلوب مجموعة كاملة من وحدات فرعية إضافية للإنتاج وظيفية الكالسيوم V 1.2 قنوات في غشاء البلازما والتفاعلات الدينامية بين هذه الوحدات الفرعية ضرورية لدعم وظيفة الكهربائية العادية للقلب 13. كاليفورنيا الخامس ß يشجع على التعبير سطح الخلية كا V 1.2 القنوات غير التساهمية nanomolar التفاعل مسعور 14. شارك في التعبير عن كا الخامس α2δ1 فرعية وايملزمة SS-تشرين الكالسيوم الخامس الكالسيوم الخامس α1 يحفز التعبير الذروة الحالية (5-10 أضعاف) ويعزز تفعيل القناة في الفولتية أكثر سلبية. الحصول على وظيفة من الطفرات من الوحيدات تشكيل المسام وقد ارتبط الكالسيوم V 1.2 مع شكل من أشكال عدم انتظام ضربات القلب البطيني تسمى متلازمة كيو تي الطويلة 15 في حين أن مجموعة من الطفرات نقطة في مفارز الرئيسية الثلاثة التي تشكل L من نوع كا V 1.2 قناة تم تحديدها في موضوعات يعانون من عدم انتظام ضربات القلب من شكل متلازمة قصيرة QT 16،17. القنوات الأيونية هي بروتينات الغشاء الذي يمكن التحقيق من وجهة نظر الكيمياء الحيوية (كيمياء البروتين) أو باستخدام أدوات الكهربية (آلات توليد الحالي) وغالبا ما تستخدم هذه المناهج التكميلية. الكهربية، ولا سيما خلية كاملة التصحيح، لقط، هو نهج مناسب لتوضيح وظيفة القنوات الأيونية 15 ولكن لا يمكن حل التعديلات في الاتجار البروتين من التغييرات في فيزيائية لهاالخصائص. الكيمياء البروتين و، ومع ذلك، وغالبا ما يقتصر استخدام بسبب التعبير منخفضة نسبيا من بروتينات الغشاء كبيرة النسبية للبروتينات قابلة للذوبان أصغر. تحتاج إلى تطوير لتعالج على وجه التحديد عيوب في نشوء حيوي البروتين يسبب تغييرات في التعبير سطح الخلية من القنوات الأيونية قوية الأساليب الإنتاجية العالية باستخدام قراءات مضان.
التدفق الخلوي هي تكنولوجيا الفيزيائية الحيوية المستخدمة في عد الخلايا، والفرز، وكشف العلامات البيولوجية، وهندسة البروتينات 18. عندما يتم حقن محلول عينة من الخلايا الحية أو الجسيمات في قياس التدفق الخلوي، يتم ترتيب الخلايا في تيار واحد التي يمكن بحثها من قبل نظام الكشف عن الجهاز (الشكل 1). تدفق الأول الكريات أداة أنتجت في عام 1956 19 الكشف عن معلمة واحدة فقط ولكن أجهزة قياس التدفق الخلوي الحديثة ليزر متعددة وأجهزة الكشف عن مضان التي تسمح للكشف عن أكثر من 30 المعلمات الفلورسنت 20،21.الفلاتر والمرايا (البصريات الانبعاث) مباشرة مبعثر خفيفة أو ضوء الفلورسنت من الخلايا لشبكة إلكترونية (الضوئي وأجهزة الكشف) التي تحول ضوء يتناسب مع شدته. ويتم تحليل البيانات الرقمية باستخدام برامج متخصصة ويتم عرض الإنتاج الأساسي باعتباره نقطة مؤامرة 21.
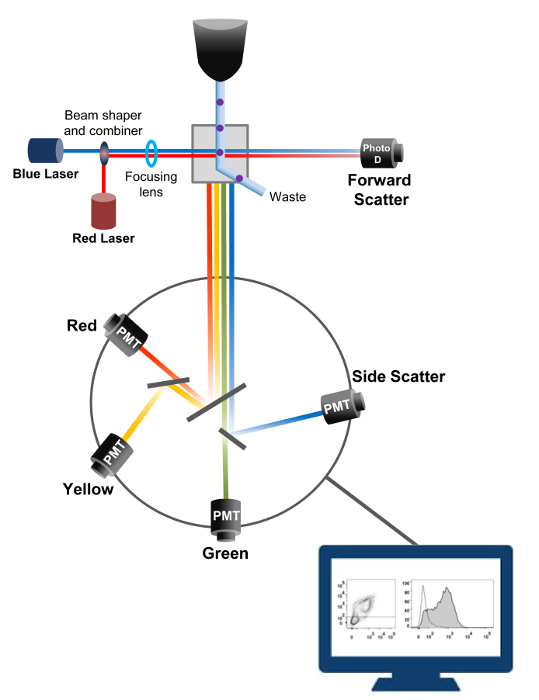
الشكل 1: مبادئ فيزيائي حيوي من تدفق الفرز الخلوي يتم دفع الخلايا واحدة من خلال فوهة تحت ضغط عال داخل تيار من السوائل غمد الذي ينقلهم عبر واحدة أو أكثر من نقاط الاستجواب الليزر. ينحرف شعاع ضوء من الخلايا تمر ويتم إرسالها ضوء جمعها في الاتجاه إلى الأمام (إلى الأمام مبعثر، FCS) إلى الثنائي الضوئي الذي يحول الضوء إلى إشارة يتناسب مع حجم الخلية. يتم جمع ضوء أيضا في زاوية 90 درجة إلى طريق الليزر وإرسالها إلى أجهزة الكشف عن (وتسمى أيضا صمام تضخيم ضوئي (PMT)).يتم توجيه هذا الضوء من خلال المرايا مزدوج اللون التي تسمح للكشف عن إشارة الجانب مبعثر (SSC)، مما يعكس تحبب داخل الخلايا، والانبعاثات الفلورسنت إذا fluorochromes متحمس موجودة في الخلية. يتم تمثيل ثلاثة أجهزة الكشف عن (الأخضر والأصفر، والأحمر) مع مرشحات مختلفة الطول الموجي ممر الموجة، والسماح للكشف في وقت واحد fluorochromes مختلفة. ورقمنة الإشارات المختلفة من خلال جهاز كمبيوتر خارجي وتحويلها إلى بيانات التي سيتم تحليلها لتحديد خصائص الخلايا. الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
وقد استغل القدرة الإنتاجية العالية من أجهزة قياس التدفق الخلوي لقياس التعبير غشاء النسبي المؤتلف من النوع البري والاتجار نقص الجهد بوابات L من نوع كا V 1.2 القنوات ومفارز المرتبطة في الخلايا الحية. كدنا] يبني المشتركوالموسومة دينغ للبروتينات مضاعفة للقيام في وقت واحد حاتمة خارج الخلية غير الفلورسنت التي يمكن الكشف عنها بواسطة الأجسام المضادة مترافق فلوري كتيمة وfluorophore الخلايا التي هي فلوري جوهري. كلا حاتمة خارج الخلية، وإدراجها في حلقة خارج الخلية من البروتين، وfluorophore داخل الخلايا، وإدراجها بعد C-محطة، وتحول مع البروتين. في هذه السلسلة من التجارب، وهندسة كا بروتين الخامس α2δ1 إبداء هيماغلوتينين خارج الخلية (HA) حاتمة (YPYDVPDYA) الكشف من قبل FITC (فلوريسئين ثيوسيانات) بردة -conjugated مكافحة HA و mCherry كما fluorophore الخلايا الذاتية. لتحديد مستوى التعبير سطح الخلية النسبي للmCherry-كا الخامس α2δ1 بروتين HA الموسومة، وخلايا تم حصادها المؤتلف التعبير عن بروتين الانصهار بعد ترنسفكأيشن، وملطخة FITC مترافق الماوس وحيدة النسيلة المضادة للHA العلامة حاتمة antibodص (الشكل 2). FITC هو مركب الفلورسنت العضوي الذي هو أصغر بكثير من صحفيين الانزيم وبالتالي ليس من المرجح لتتداخل مع وظيفة بيولوجية. mCherry- الكالسيوم الخامس α2δ1-HA overexpressed في TSA-201cells، وتنتج عن زيادة قدرها 3-سجل كبيرة في مضان FITC وmCherry مضان على قطع ثنائية الأبعاد 22. وبالنظر إلى أن حاتمة HA يقع في جزء الخلية من البروتين، وكثافة مضان لFITC التي تم الحصول عليها في وجود خلايا سليمة تعكس المؤشر النسبي للتعبير عن سطح الخلية من البروتين HA الموسومة. إمكانية الوصول إلى حاتمة HA في بنيات يتم التحقق بشكل منهجي عن طريق قياس إشارة FITC بعد permeabilization الخلية. يخدم هذا الإجراء أيضا لإثبات مجموع تعبير البروتين تطبيع منذ كثافات مضان النسبية لFITC قدرت في الخلايا permeabilized قابلة للمقارنة نوعيا إلى القيم النسبية مضان FOص mCherry تقاس في ظل ظروف permeabilized وغير permeabilized 22،23. من المهم أن نلاحظ أن الطيف مضان جوهري هو تحول نحو القيم العليا بعد permeabilization إلا أن القيمة الوحيدة التي أبلغ عنها هو التغير في كثافة مضان بالمقارنة مع بناء السيطرة. وتقدر التغيرات النسبية في كثافة مضان لبنيات الاختبار باستخدام كثافة ΔMean الإسفار (ΔMFI) القيم لكل fluorophore (mCherry أو FITC). تم تصميم التجارب لقياس كثافة مضان الاختبار بناء بالنسبة للكثافة مضان من بناء سيطرة أعرب تحت نفس الظروف للحد من الاختلافات التجريبية في مضان لا يتجزأ من الأجسام المضادة fluorophore مترافق. درست اثنين من البروتينات الغشاء بنجاح باستخدام هذا الاختبار: الوحيدات تشكيل المسام من L-نوع الجهد بوابات قنوات الكالسيوم الكالسيوم V 1.2 14،22 وفي سلسلة مختلفة منالتجارب، مساعد خارج الخلية كا الخامس α2δ1 فرعية 22،23. تم استخدام بروتوكول التالية لتحديد التعبير سطح الخلية من الكالسيوم الخامس α2δ1 فرعية من القلب L من نوع كا V 1.2 قناة ظل ظروف السيطرة وبعد الطفرات التي تؤثر على تعديل posttranslational من القناة الايونية. تحت ظروف تجريبية موحدة، ومضان سطح الخلية من FITC يزيد شبه خطيا مع التعبير عن كدنا] الترميز لmCherry-كا الخامس البروتينات α2δ1-HA (الشكل 5 من مرجع 22).
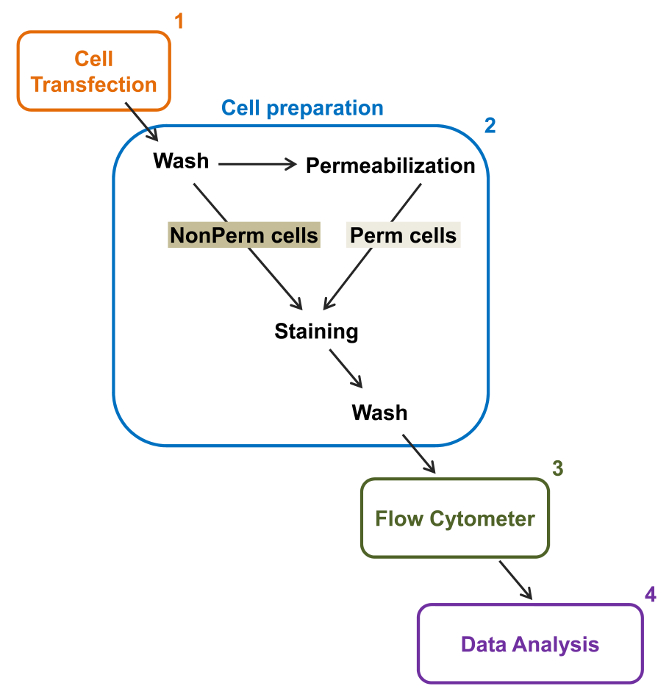
الشكل 2: تمثيل تخطيطي لوضع العلامات الكلي والغشاء في التدفق الخلوي بروتوكول تجريبي ويبين المخطط بعض الخطوات الرئيسية اللازمة لقياس نسبي تعبير كامل وسطح الخلية من القنوات الأيونية المؤتلف التي كتبها فلوريداآه الخلوي. و transfected الخلايا مع المزدوج معلم البناء mCherry-كا الخامس α2δ1-HA في TSA-201 الخلايا (1) وملطخة قبل أو بعد permeabilization (2). يتم الحصول على البيانات Multiparameter في قياس التدفق الخلوي (3) للتحليل متعدد المتغيرات (4). الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
