Dieses Dokument bietet einen zuverlässigen Assay, um die relative Zelloberflächenexpression der Membranproteine wie Ionenkanäle exprimiert in rekombinanten Zellen unter Verwendung der bestehenden Technologie Durchflusszytometrie zu melden. Ionenkanäle sind porenbildende Membranproteine, die durch Gating den Fluss von Ionen durch die Zellmembran zum Steuern elektrischer Signale verantwortlich sind. Sie werden durch den Aktivierungsmechanismus, der Natur klassifiziert und Selektivität der Ionenspezies durch die Pore queren, wo sie lokalisiert sind. Auf zellulärer und Gewebespiegel, die makroskopischen Ionenflüsse durch Ionenkanäle sind das Produkt von biophysikalischen (Gating und Permeation), biochemische (Phosphorylierung) und Biogenese (Synthese, Glykosylierung, Menschenhandel und Abbau) Eigenschaften 1. Jedes dieser Verfahren ist einzigartig für jede Art von Ionenkanälen und optimiert wird, um die physiologische Rolle des Ionenkanals zu erfüllen. Folglich Veränderungen in jedem dieser fein abgestimmten Prozesse durch einevererbte oder eine genetische Modifikation, die oft als "Ionenkanal" kann zu Zellhomöostase schädlich sein. Es ist wichtig zu betonen, dass an der Zelloberfläche, die "richtige" Menge an Ionenkanälen liefern zu Zellhomöostase kritisch ist. Selbst kleine Erhöhungen (Gain-of-function) und geringe Abnahmen (loss-of-function) in Ionenkanalaktivität haben das Potenzial, eine ernsthafte Pathologie im Laufe eines Lebens zu verursachen. Defekte in der Zelloberfläche Lieferung von reifen Ionenkanäle ist ein wichtiger Faktor in zahlreichen channelopathies, wie zystischer Fibrose (CFTR Ionenkanal) 2 und Herzrhythmusstörungen des Long – QT – Syndrom Form (kardiale Kaliumkanäle) 3.
Kanalopathien sind mit Herz plötzlichen Tod 4 verbunden. Die aktuelle weltweite Prävalenz aller Herz channelopathies wird angenommen , mindestens 1: 2,000-1: etwa die Hälfte des plötzlichen Herztods arrhythmic ca. 3.000 pro Einzel 5 und sind verantwortlichses 6. Dysfunction in Herzspannungsabhängigen Natrium-, Kalium- und kalzium selektive Ionenkanäle sind dafür bekannt, eine Schlüsselrolle in diesem Prozess spielen. Der L-Typ – Ca V 1.2 spannungsabhängigen Calcium – Kanal erforderlich synchronisiert Herzmuskelkontraktion zu initiieren. Der Herz L-Typ – Ca V 1.2 – Kanal ist ein Multi-Untereinheiten – Proteinkomplex , bestehend aus dem Hauptporenbildenden Ca V α1 – Untereinheit und Ca V SS und V Ca α2δ1 Hilfsuntereinheiten 7-12. Beachten Sie, dass die volle Ergänzung der Hilfsuntereinheiten funktionellen Ca V 1.2 Kanäle an der Plasmamembran und dynamische Wechselwirkungen zu erzeugen erforderlich ist zwischen diese Untereinheiten sind wichtig , die normale elektrische Funktion des Herzens 13 zu unterstützen. Ca V ß fördert die Zelloberflächenexpression von Ca V 1.2 Kanäle , durch eine nicht-kovalente nanomolar hydrophobe Interaktionschromatographie 14. Die Co-Expression des Ca V α2δ1 Untereinheit with Ca V ß-gebundene Ca V α1 stimuliert Spitzenstrom Ausdruck (5 bis 10-fach) und fördert Kanalaktivierung bei negativeren Spannungen. Gain-of-function Mutationen des porenbildende Untereinheit Ca V 1.2 haben mit einer Form von ventrikulären Arrhythmien sogenannte Long – QT – Syndrom 15 , während eine Vielzahl von Punktmutationen in den drei Hauptuntereinheiten bilden , die L-Typ – Ca V 1.2 Kanal zugeordnet worden wurden bei Patienten leiden unter Arrhythmien des Short – QT – Syndrom Form 16,17 identifiziert. Ionenkanäle sind Membranproteine, die aus biochemischer Sicht (Proteinchemie) oder mittels elektro Tools (Stromerzeugungsmaschinen) und häufig diese komplementären Ansätzen untersucht werden können. Electrophysiology, insbesondere Ganzzell – Patch-Clamp ist ein geeigneter Ansatz , um die Funktion von Ionenkanälen 15 zu erläutern , aber nicht lösen können Modifikationen in den Proteintransport durch Veränderungen in ihren biophysikalischenEigenschaften. Proteinchemie hat jedoch häufig den Einsatz aufgrund der relativ geringen Expression großer Membranproteine in Bezug auf kleinere lösliche Proteine beschränkt. Robuste Hochdurchsatzverfahren Fluoreszenzablesung unter Verwendung müssen entwickelt werden, um gezielt Defekte in Proteinbiogenese adressieren Veränderungen in der Zelloberflächenexpression von Ionenkanälen verursacht.
Die Durchflusszytometrie ist ein biophysikalischen Technologie in Zellzählung eingesetzt, das Sortieren, den Nachweis von Biomarkern und Protein – Engineering – 18. Wenn eine Probenlösung von lebenden Zellen oder Partikel in einem Durchflusszytometer eingespritzt wird, werden die Zellen in einen einzigen Strom bestellt, die von der Maschine des Detektionssystems (Figur 1) untersucht werden können. Die erste Durchflusszytometer Instrument 1956 erzeugt 19 detektiert nur einen Parameter , aber moderne Durchflußzytometer weisen mehrere Laser und Fluoreszenzdetektoren, die den Nachweis von mehr als 30 Fluoreszenzparameter 20,21 ermöglichen.Filter und Spiegel (Emissionsoptik) lenken das Licht streuen oder Fluoreszenzlicht von Zellen an ein elektronisches Netzwerk (Photodiode und Detektoren), die das Licht proportional zu seiner Intensität umwandeln. Digitale Daten werden mit spezieller Software analysiert und die primäre Ausgabe wird als Punkt – Diagramm 21 angezeigt.
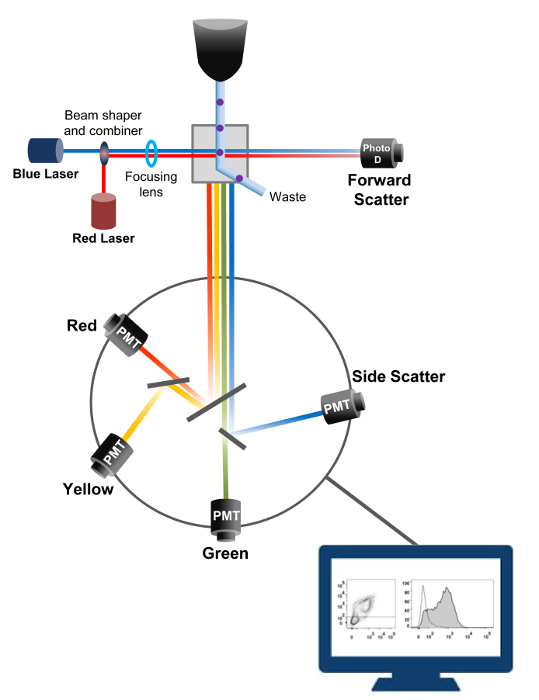
Abb . 1: Biophysical Prinzipien der Durchflusszytometrie Sortieren einzelner Zellen werden durch eine Düse unter hohem Druck in einem Strom von Hüllflüssigkeit geschoben , der sie bewegt sich über eine oder mehrere Laserabfragepunkten. Der Lichtstrahl wird durch die vorbeifahrenden Zellen umgelenkt und das Licht in der Vorwärtsrichtung gesammelt (Forward Scatter, FCS) auf eine Fotodiode geschickt, die das Licht in ein Signal proportional zur Größe der Zelle umwandelt. Das Licht wird auch in einem 90 ° Winkel zu dem Laserpfad und an Detektoren (auch als Photomultiplier (PMT)) gesammelt.Dieses Licht wird durch den dichroitischen Spiegel geleitet, die die Erkennung des Seitenstreusignal (SSC) zu ermöglichen, die die Granularität innerhalb der Zellen widerspiegelt, und die Fluoreszenzemissionen, wenn erregt Fluorochrome in der Zelle vorhanden sind. Drei Detektoren (grün, gelb und rot) mit unterschiedlichen Wellenlängen-Bandpassfilter dargestellt, so dass die gleichzeitige Detektion von verschiedenen Fluorochromen. Die unterschiedlichen Signale werden von einem externen Computer digitalisiert und in Daten umgewandelt, die die Eigenschaften der Zellen zu quantifizieren analysiert werden. Bitte hier klicken , um eine größere Version dieser Figur zu sehen.
Die Hochdurchsatzkapazität von Durchflusszytometer wurde die relative Membranexpression von rekombinanten Wildtyp und des Handels-defizienten spannungsabhängigen L-Typ – Ca V 1.2 Kanäle und die damit verbundenen Untereinheiten in lebenden Zellen zu quantifizieren ausgebeutet. cDNA-Konstrukten coding für die Proteine wurden doppelt markiert, um gleichzeitig eine extrazelluläre nichtfluoreszierenden Epitop tragen, die durch eine undurchlässige fluoreszierenden konjugierten Antikörper und einem intrazellulären Fluorophor die konstitutiv fluoresziert detektiert werden kann. Sowohl das extrazelluläre Epitop in einer extrazellulären Schleife des Proteins eingeführt ist, und die intrazelluläre Fluorophor, eingesetzt nach dem C-Terminus mit dem Protein translatiert. In dieser Serie von Experimenten wurde die Ca V α2δ1 proteintechnisch eine extrazelluläre Hämagglutinin (HA) -Epitop (YPYDVPDYA) durch eine undurchlässige FITC (Fluorescein – Isothiocyanat) -konjugiertem anti-HA und mCherry als Intrinsische intrazellulärem Fluorophor detektiert auszudrücken. Um die relative Zelloberflächenexpressionsniveau des mCherry-Ca V α2δ1 HA-markiertes Protein, rekombinante Zellen, die das Fusionsprotein bestimmen , wurden nach der Transfektion geerntet, und gefärbt mit FITC-konjugierten monoklonalen Maus – anti-HA – Epitop – Tag Antibody (Abbildung 2). FITC ist eine organische Fluoreszenzverbindung, die wesentlich kleiner als Enzym-Reporter ist und deshalb nicht so wahrscheinlich mit biologischen Funktion zu stören. mCherry- Ca V α2δ1-HA überexprimiert in Tsa-201cells, erzeugt eine signifikante 3-log Anstieg der FITC – Fluoreszenz und mCherry Fluoreszenz auf zweidimensionalen Plots 22. Da die HA-Epitop in der extrazellulären Teil des Proteins befindet, erhalten die Fluoreszenzintensität für FITC in Gegenwart von intakten Zellen spiegeln die relativen Index der Zelloberflächenexpression von HA-markierten Proteins. Die Zugänglichkeit des HA-Epitops in den Konstrukten wird durch Messung des FITC Signals nach Zellpermeabilisierung systematisch validiert. Auch diese Maßnahme dient der normalisierten Gesamtproteinexpression zu untermauern, da die relativen Fluoreszenzintensitäten für FITC in permeabilisierten Zellen geschätzt sind qualitativ vergleichbar mit den relativen Fluoreszenzwerte for mCherry gemessen unter permeabilisiert und nicht-permeabilisierten Bedingungen 22,23. Es ist wichtig zu beachten, dass die intrinsische Fluoreszenzspektrum zu höheren Werten nach Permeabilisierung verschoben wird, sondern daß der einzige Wert angegeben wird, ist die Änderung in der Fluoreszenzintensität in Bezug auf die Kontrollkonstrukt verglichen. Relative Änderungen in der Fluoreszenzintensität für die Testkonstrukte werden geschätzt die ΔMean Fluoreszenzintensität unter Verwendung von (ΔMFI) Werte für jedes Fluorophor (mCherry oder FITC). Experimente werden die Fluoreszenzintensität des Testkonstrukts relativ zur Fluoreszenzintensität des Kontrollkonstrukts unter den gleichen Bedingungen exprimiert zu messen experimentellen Variationen in der intrinsischen Fluoreszenz des Fluorophors-konjugierten Antikörper zu begrenzen. Zwei Membranproteine wurden erfolgreich mit diesem Test untersucht: das porenbildende Untereinheit des L-Typ spannungsabhängigen Calcium – Kanal – Ca V 1.2 14,22 und in einer anderen Reihe vonExperimente, 22,23 die extrazelluläre Hilfs Ca V α2δ1 Untereinheit. Das folgende Protokoll wurde verwendet , um die Zelloberflächenexpression des Ca V α2δ1 Untereinheit des Herz L-Typ – Ca V 1.2 Kanal unter Kontrollbedingungen und nach Mutationen zu bestimmen , die posttranslationale Modifikation des Ionenkanals beeinflussen. Unter genormten Versuchsbedingungen erhöht die Zelloberfläche Fluoreszenz von FITC quasi-linear mit der Expression der cDNA kodierend für die mCherry-Ca V α2δ1-HA – Proteine (Abbildung 5 aus Lit. 22).
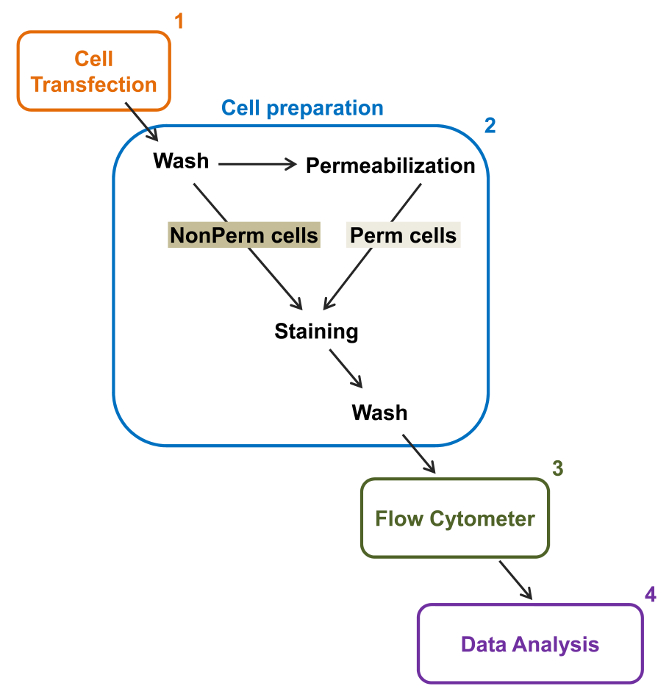
Abbildung 2:. Schematische Darstellung der gesamten und der Membran Kennzeichnung in der Durchflusszytometrie Versuchsprotokoll Das Schema beschreibt einige der wichtigsten Schritte notwendig , um die relativen Gesamt und Zelloberflächenexpression von rekombinanten Ionenkanäle durch fl zu quantifizierenow-Zytometrie. Zellen werden mit dem doppelt markiert Konstruktion mCherry-Ca V α2δ1-HA in TSA-201 – Zellen transfiziert (1) und gefärbt vor oder nach der Permeabilisierung (2). Multi Daten werden in einem Durchflusszytometer (3) für multivariate Analyse (4) übernommen. Bitte hier klicken , um eine größere Version dieser Figur zu sehen.
