Analyse de la croissance dépend d'un ensemble d'outils qui sont couramment utilisés par les scientifiques de plantes pour décrire le génotype déterminé les différences de croissance et / ou des réponses phénotypiques à des facteurs environnementaux. Ils comprennent la taille et le poids des mesures de la plante entière ou un organe et les calculs des taux de croissance à explorer les mécanismes sous-jacents de la croissance. Croissance organique est déterminée par la division cellulaire et de l'expansion au niveau cellulaire. Par conséquent, incluant la quantification de ces deux procédés dans des analyses de croissance est essentiel pour comprendre les différences de croissance organe entier 1. Par conséquent, il est crucial d'avoir une méthodologie appropriée pour déterminer les paramètres de croissance cellulaire qui est relativement facile à utiliser par des laboratoires non spécialisés.
L' analyse cinématique a déjà été établie comme une approche fournissant un cadre puissant pour le développement de modèles de croissance de l' organe 2. La technique a été optimisé pour les systèmes linéaires,tels que les racines d' Arabidopsis thaliana et les feuilles monocotylédones, mais également pour les systèmes non-linéaires, tels que les feuilles dicotylédone 3. De nos jours, cette méthodologie est de plus en plus utilisée pour étudier comment génétiques, hormonaux, développementaux et les facteurs environnementaux influencent la division cellulaire et l' expansion dans divers organes (tableau 1). En outre, il fournit également un cadre pour lier les processus cellulaires à leurs règlements biochimiques, moléculaires et physiologiques sous – jacents (tableau 2), bien que les limites peuvent être imposées par la taille des organes et de l' organisation spatiale des techniques qui nécessitent des quantités plus élevées de matières végétales (par exemple, métabolite mesures, de la protéomique, etc.).
Feuilles monocotylédones telles que le maïs (Zea mays) feuille, représentent des systèmes linéaires dans lesquels les cellules se déplacent à partir de la base de la feuille vers la pointe, en passant successivement à travers la zone de méristèmes et de l' allongement pour atteindre la maturitézone. Cela en fait un système de modèle idéal pour les études quantitatives de la configuration spatiale de la croissance 4. De plus, les feuilles de maïs ont de grandes zones de croissance (méristème et zone d'allongement couvrant plusieurs centimètres 5) et offrent des possibilités d'études à d' autres niveaux d' organisation. Cela permet de l'enquête sur les mécanismes de régulation (putatifs) contrôlant la division cellulaire et de l' expansion, quantifiée par analyse cinématique grâce à une gamme de techniques moléculaires, des mesures physiologiques, et des approches de biologie cellulaire (tableau 2).
Ici, nous fournissons un protocole pour effectuer une analyse cinématique dans les feuilles monocotylédones. Tout d'abord, nous expliquons comment procéder à une analyse appropriée à la fois la division cellulaire et l'élongation des cellules en fonction de la position le long de l'axe de la feuille et la façon de calculer les paramètres cinématiques. Deuxièmement, nous montrons aussi comment cela peut être utilisé comme base pour la conception d'échantillonnage. Ici, nous discutons deux cas: haute résolution d'échantillonnage d'und axée échantillonnage, permettant une meilleure interprétation des données et le gain de temps / argent, respectivement.
Tableau 1. Aperçu des analyses cinématique des méthodes de quantification de la division cellulaire et de l' expansion dans divers organes.
| organe | référence |
| feuilles monocotylédones | 16, 20, 21, 22 |
| pointes des racines | 2, 23, 24, 25, 26, 27, 28, 29 |
| feuilles dicotylédones | 21, 30, 31 |
| méristème apical | 32 |
Tableau 1. Aperçu des analyses cinématique des méthodes de quantification de la division cellulaire et de l' expansion dans divers organes.
<p class="jove_content" fo:keep-together.within-page = "1">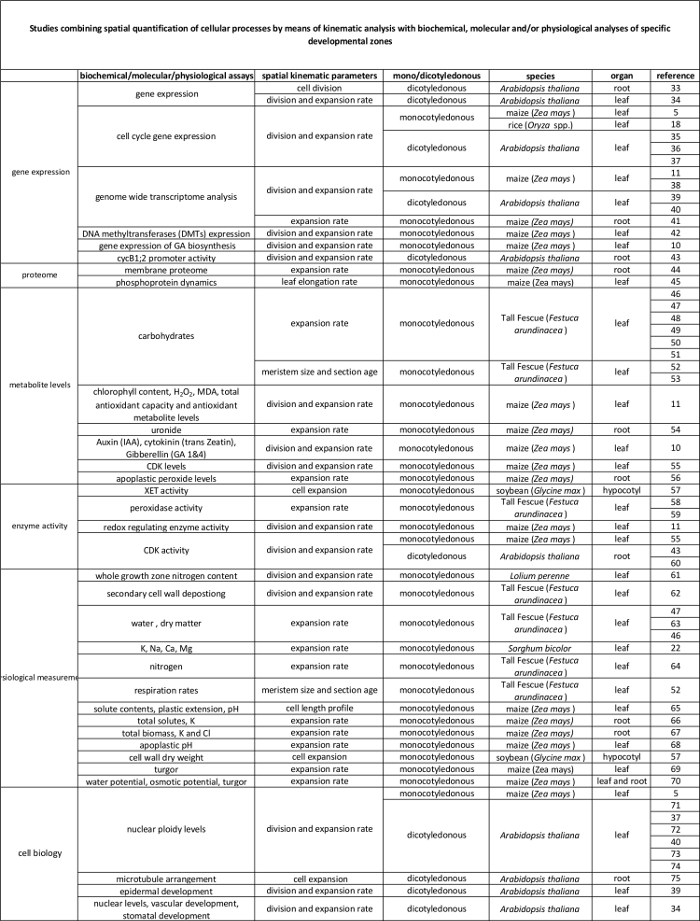
Tableau 2. Lien entre les processus cellulaires quantifiés par l'analyse cinématique à leur régulation au niveau moléculaire. Les références à diverses études reliant la quantification des processus cellulaires aux résultats des tests biochimiques et moléculaires chez diverses espèces et des organes. Endotransglucosylase Xyloglucan (XET), malondialdéhyde (MDA), les kinases cycline-dépendantes (CDK). S'il vous plaît cliquer ici pour voir une version plus grande de cette table.
