De dissociatieconstante K is een belangrijke parameter voor het beschrijven van de affiniteit van een receptor (R) voor haar ligand (L). Op basis van de wet van massa-actie, wordt K gedefinieerd voor het evenwicht, waarin de receptor-ligand complexe RL in de receptor R en de ligand L: dissocieert
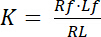 Vergelijking 1
Vergelijking 1
met de indexcijfers f de gratis of ongebonden status van receptor en ligand. De concentratie van het complex van receptor-ligand, RL, is gelijk aan de concentratie van de receptor-gebonden ligand Lb. Als de totale concentratie van receptor Rt de som van de vrije receptor Rf en ligand-gebonden receptor Rb is = Lb, de dissociatieconstante kan ook geschreven worden als:
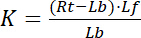 Vergelijking 2
Vergelijking 2
Dus, de verzadiging opbrengst Y, gedefinieerd als de breuk van afhankelijke ligand Lb ten opzichte van de totale concentratie van receptor Rt,
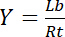 Vergelijking 3
Vergelijking 3
afhankelijk van de concentratie van gratis ligand Lf:
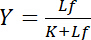 Vergelijking 4
Vergelijking 4
Deze hyperbolische relatie beschrijft de bindende curve van een receptor-ligand interactie en zijn plot toont de concentratie van afhankelijke ligand Lb als een functie van de concentratie van gratis ligand Lf. Van de bindende curve, kan de dissociatieconstante K worden afgeleid als de concentratie van gratis ligand op half-maximale verzadiging opbrengst. Bovendien zijn verschillende algoritmen linearize bindende curven vastgesteld, zoals de dubbele-wederkerige plot door Klotz1,2, of transformaties volgens Scatchard of Hanes (herzien door Bisswanger3). Maar lijdt alle algoritmen aan het probleem dat de maximale waarde van de verzadiging opbrengst, die asymptotisch bij hoge concentraties van gratis ligand in de bindende curve wordt benaderd, moet worden geraamd in een grafische pre evaluatie en daarom is vergissing-geneigd.
Bovendien, vereist de vaststelling van een bindende curve de kwantificering van vrije en gebonden ligand tijdens het evenwicht bindend. Te dien einde moet de gratis ligand worden gescheiden van het ligand-receptor-gebonden en gekwantificeerd. Daarom moeten de ligand en receptor verschillen in hun eigenschappen, zoals een niet-protein ligand in tegenstelling tot een eiwit-receptor. Als beide partners bindend eiwitten zijn, moeten zij worden onderscheiden in hun formaat, kosten of andere moleculaire kenmerken. De kwantificering van de concentraties van het ligand in kleinschalige vrijblijvende aanpak is echter een moeilijke taak. Radioactieve labeling van het ligand behoefde vaak te detecteren de lage concentratie van afhankelijke ligand, vooral als aanzienlijke bedragen van receptoren niet beschikbaar of betaalbaar waren. Bovendien kan de receptor-gebonden ligand tijdens en na de isolatie op een niet te verwaarlozen wijze distantiëren. Vandaar, complexe methoden, zoals evenwicht gel filtratie4, capillaire elektroforese5en pulse proteolyse6, zijn vereist voor het kwantificeren van receptor-gebonden ligand en scheiden van gratis ligand.
In tegenstelling tot deze bindende analyses vereisen titratie experimenten geen kwantitatieve scheiding van afhankelijke en gratis ligand. Te dien einde wordt een receptor op een constante concentratie getitreerd met verschillende concentraties van toegevoegde ligand. Door binding aan de receptor heeft de afhankelijke ligand een biofysische eigenschap die zich onderscheidt van de gratis ligand en meetbaar door, bijvoorbeeld, fotometrie, fluorometry of antistofdetectie. Dus, een signaal van S, die evenredig is met de verzadiging opbrengst Y en bijgevolg ook de concentratie van receptor-gebonden ligand (L-b), wordt gedetecteerd als een functie van de totale concentratie van toegevoegde ligand (Lt). Parameters, het signaal S zowel de totale concentratie van toegevoegde ligand zijn gekwantificeerd op een directe en gemakkelijker manier dan de concentraties van afhankelijke en gratis ligand. Vooral het opsporen van receptor-gebonden ligand door enzyme-linked immunosorbent assay (ELISA) toegestaan de vermindering van de steekproef volumes tot onder 100 µL en parallelle metingen van verschillende concentraties van de ligand in multi goed microtiterplaat platen. In een titratie ELISA, is een receptor fysiek geadsorbeerd aan een plaat van de microtiterplaat op dezelfde concentratie en getitreerd met oplosbare ligand. De receptor is in wezen door hydrofobe adsorptie aan het plastic oppervlak geïmmobiliseerd. De oppervlakte concentratie van geïmmobiliseerdet receptor correlaten met de concentratie van de coating van de receptor in een niet-lineaire relatie, waarschijnlijk volgens Langmuir´s adsorptie] Thermo7. Naast het totale aantal geadsorbeerde receptor moleculen is hun activiteit staat een andere belangrijke parameter voor titratie testen. Alleen geïmmobiliseerd receptoren die ligand bindende activiteit hebben behouden, relevant zijn voor de bepaling van de titratie en uiteindelijk bijdragen aan de totale concentratie aan actieve receptoren Rt van de titratie assay, die direct kan niet worden bepaald.
Sites op het plastic oppervlak, die niet onder de geïmmobiliseerdet receptor vallen zijn gevoelig voor andere eiwitten, zoals de ligand adsorberen. Fysische adsorptie van het ligand dergelijke kunststof oppervlak sites zou resulteren in een vergelijkbaar signaal als het ligand-receptor-gebonden, maar toch in een niet-specifieke wijze. Om dit niet-specifieke signaal, het plastic oppervlak sites van de microtiterplaat platen die niet hebben zijn bekleed met eiwit nog wordt geblokkeerd met bovien serumalbumine (BSA). Echter voor sommige receptor-ligand titratie testen, kunnen niet-specifieke achtergrond signalen worden waargenomen. Dan, andere blokkerende makelaars, zoals een oplossing van 0,2% gelatine of van 0,04% Tween 20, worden aanbevolen.
Na binding aan de receptor, wordt de gratis ligand verwijderd door twee wassen stappen. Afhankelijke ligand blijft met de receptor, dat is geïmmobiliseerd tot het plastic oppervlak van de microtiterplaat put en eventueel versterkt door chemische fixatie. Voor de latere covalente Kruis-koppeling van afhankelijke ligand en geïmmobiliseerdet receptor met Glutaaraldehyde, wordt de buffer stof TRIS vervangen voor HEPES, zonder enige verandering in de ligand bindende. HEPES, in tegenstelling tot TRIS, doet niet inactivering van Glutaaraldehyde. De covalente Kruis-koppeling met Glutaaraldehyde corrigeert de afhankelijke ligand met de receptor en voorkomt dat de dissociatie tijdens opeenvolgende wassen en incubatie stappen. Dus, de receptor-ligand interactie is chemisch bevroren en garandeert een titratiecurve die beïnvloed door de opeenvolgende stappen van wassen en incubatie wordt. Glutaaraldehyde fixatie kan echter chemisch wijzigen de ligand en receptor op zodanige wijze dat hun interactie is verlaagd of afgeschaft. Bovendien, wijziging van epitopes binnen de ligand kan veranderen de bindende affiniteit van het antilichaam detecteren, vooral als een monoclonal antilichaam wordt gebruikt om te kwantificeren afhankelijke ligand. Hoewel geen van deze negatieve effecten van Glutaaraldehyde fixatie in deze titratie ELISA optreedt, moet de gevoeligheid van de test naar Glutaaraldehyde elke receptor-ligand interactie vóór de titratie experiment worden getest. Na fixatie, overtollige Glutaaraldehyde verwijderd in drie stappen van het wassen met TRIS-bevattende buffer. TRIS inactiveert overige aldehyde-groepen, die nonspecifically reageren misschien met het opsporen van antilichamen in de volgende stap.
Het bedrag van de afhankelijke ligand wordt gekwantificeerd met enzym-verbonden antilichamen, waarmee een fotometrische ELISA signaal S. Dit is uitgezet tegen de totale ligand concentratie Lt toegevoegd aan elk putje. Ondanks zijn gemakkelijker aanwinst is de titratiecurve niet een hyperbolische functie in tegenstelling tot de bindende curve. Bovendien is onduidelijk hoe de berekening van de dissociatieconstante K uit een titratiecurve. Hoewel algoritmen linearize spectroscopically verworven titratie curven zijn onafhankelijk gemeld door Stockell8 en Heyn en Weischet9, ze viel kort vanwege hun onzekerheid van de schatten van het signaal maximale waarde die de verzadiging opbrengst benaderingen bij hoge concentraties van toegevoegde ligand.
Hier, zijn een titratie ELISA en een niet-lineaire regressie-algoritme beschreven voor het afleiden van de dissociatieconstante K voor een receptor-ligand interactie van een titratiecurve. Dit protocol wordt geïllustreerd voor de interactie van het collageen-bindende A-domein van het integrine-α2β1 met een slang GIF-afgeleide-remmer. Verschijnsel zijn cel celadhesie-moleculen, die de ankerplaats van cellen aan de omliggende extracellulaire matrix of de onderliggende kelder membraan10,11 bemiddelen. Bovendien overbrengen verschijnsel belangrijke signalen tussen cellen en de extracellulaire matrix door de indienstneming van extra signalering van moleculen en de vorming van nieuwe organellen van de cel, adhesomes, op cel-matrix interactie12,13, 14. collageen, de ligand van integrine α2β1, is de meest voorkomende eiwit van het menselijk lichaam en is een onderdeel van de cruciale steigers van het bindweefsel15. De interactie tussen α2β1 integrine en collageen wordt gemedieerd door de A-domein van de integrine alpha2 subeenheid. Het integrine α2A-domein bevat een divalente kationen, die is vereist voor collageen bindende en stabiliseert de structuur. De wild-type formulier, evenals de mutanten van het domein van de α2A, zoals die waarin het residu van oppervlak-blootgesteld Y216 was vervangen voor een glycine, kunnen gemakkelijk worden recombinantly in een bacteriële expressie systemen geproduceerd en geïsoleerd via hun oligo-zijne-tags met een NiNTA superflow kolom met een latere dialyse tegen TRIS-gebufferde zoutoplossing (TBS; 50 mM TRIS/HCl, pH 7.4, 150 mM NaCl) met 2 mM MgCl216. De concentraties ervan werden vastgesteld met de bicinchoninic zure test (BCA) en hun purities zijn getest door conventionele SDS-PAGE en gekleurd met Coomassie-Brilliant Blue R250.
De interactie tussen α2β1 integrine en collageen wordt geblokkeerd door de binding van de snake venom component, rhodocetin, van de Malayan pit-viper (Calloselasma rhodostoma)16,17. Gebruikt als een oplosbare ligand in deze titratie ELISA, rhodocetin was gezuiverd van de ruwe venom zoals eerder beschreven16. Het wordt ontbonden in HEPES-gebufferde zoutoplossing (HBS; 10 mM HEPES/NaOH, pH 7.4, 150 mM NaCl) en worden opgeslagen bevroren bij-20 ° C. De concentratie ervan werd bepaald door BCA en haar zuiverheid werd bewezen door SDS-PAGE. Als een antagonist, rhodocetin niet alleen geblokkeerd collageen te binden aan het integrine α2β1 A-domein, maar ook de inactieve conformatie van het integrine waardoor elke signalering van collageen in cellen of trombocyten18stabiliseert. Het is van groot belang is voor het bepalen van de dissociatieconstante van rhodocetin met haar doelstelling van receptor en dus het ontrafelen van het moleculaire mechanisme en farmaceutische potentiële bvals antitrombotica agent biomedische. Te dien einde een titratie ELISA wordt beschreven, met inbegrip van de evaluatie, die is van toepassing op bijna elke receptor-ligand interactie met een 1:1 interactie stoichiometrie.
