De mogelijkheid om ingenieur van het genoom van elk levend organisme heeft vele biomedische en biotechnologische toepassingen, zoals de correctie van de ziekte-veroorzakende mutaties, bouw van accurate cellulaire modellen voor ziekte studies of generatie van landbouw gewassen met wenselijke eigenschappen. Sinds het begin van de eeuw, verschillende technologieën zijn ontwikkeld voor genoom engineering in zoogdiercellen, met inbegrip van meganucleases1,2,3, elektrolytisch vinger nucleasen4,5, of transcriptie activator-achtige effector nucleasen (TALENs)6,7,8,9. Echter zijn deze eerdere technologieën moeilijk te programma of vervelend te monteren, waardoor hun wijdverspreide goedkeuring in onderzoek en de industrie te belemmeren.
In de afgelopen jaren de geclusterde regelmatig interspaced korte palindromische herhaalt (CRISPR) – CRISPR-geassocieerde (Cas) systeem heeft ontpopt als een krachtige nieuwe genoom engineering technologie10,11. Oorspronkelijk een adaptieve immuunsysteem in bacteriën, is met succes ingezet voor het wijzigen van het genoom in planten en dieren, inclusief de mens. Een primaire reden waarom CRISPR-Cas heeft populariteit zo veel in zo’n korte tijd is dat het element dat de belangrijkste Cas-endonuclease, zoals Cas9 of Cas12a brengt (ook bekend als Cpf1), naar de juiste locatie in het genoom is gewoon een klein stukje chimeer één gids RN A (sgRNA), die is ontwerp eenvoudig en goedkoop te synthetiseren. Na wordt gerekruteerd om de doelsite, het Cas-enzym functioneert als een paar van moleculaire schaar en cleaves het afhankelijke DNA met haar RuvC, HNH of NOC domeinen12,13,14. De resulterende dubbele gestrande onderbreking (DSB) wordt later hersteld door de cellen via niet-homologe einde deelname aan (NHEJ) of de homologie geleide reparatie (HDR) traject. Bij gebrek aan een reparatie-sjabloon, wordt de DSB hersteld door de vergissing-geneigd NHEJ traject, die aanleiding tot pseudo-willekeurige invoeging of schrapping van nucleotiden (microdeleties) op de gesneden site geven kan, mogelijk veroorzaakt frameshift mutaties in eiwit-codeert genen. Echter in de aanwezigheid van een donor-sjabloon die de gewenste DNA wijzigingen bevat, wordt de DSB hersteld door het traject van de HDR high fidelity. Gangbare typen van donor sjablonen zijn single-stranded oligonucleotides (ssODNs) en plasmiden. De voormalige wordt meestal gebruikt als de beoogde DNA veranderingen klein (bijvoorbeeld wijziging van een enkelvoudige basenpaar), zijn terwijl de laatste meestal gebruikt wordt als men het wil invoegen een relatief lange reeks (bijvoorbeeld de codering opeenvolging van een groen fluorescent proteïne of GFP) in de locus van het doel.
De endonuclease activiteit van het eiwit van Cas vereist de aanwezigheid van een protospacer aangrenzende motief (PAM) op de target website15. De PAM van Cas9 is aan de 3′-eind van de protospacer, terwijl de PAM van Cas12a (ook wel Cpf1 genoemd) in plaats daarvan aan het einde 5′16. Het Cas-guide RNA complex is niet in staat om een DSB als de PAM afwezig17 is. Vandaar, de PAM legt een beperking op de genomic locaties waar een bepaalde Cas nuclease vermag klieven. Gelukkig, Cas nucleasen van verschillende bacteriesoorten vertonen meestal verschillende PAM eisen. Vandaar, door de integratie van verschillende systemen van de CRISPR-Cas in onze engineering toolbox, we kunnen uit te breiden het bereik van sites die in een genoom kunnen worden gericht. Bovendien kan een natuurlijke Cas enzym worden ontworpen of geëvolueerd om te herkennen van de alternatieve opeenvolgingen van het PAM, verdere uitbreiding van het toepassingsgebied van genomic doelstellingen toegankelijk voor manipulatie18,19,20.
Hoewel meerdere CRISPR-Cas systemen beschikbaar voor technische doeleinden genoom zijn, hebben de meeste gebruikers van de technologie vertrouwd voornamelijk op de Cas9 nuclease van Streptococcus pyogenes (SpCas9) om meerdere redenen. Ten eerste, het vereist een relatief eenvoudig NGG PAM, in tegenstelling tot veel andere certificeringsinstanties eiwitten die kunnen alleen in de aanwezigheid van meer complexe PAMs klieven. Ten tweede, het is de eerste Cas endonuclease om te worden geïmplementeerd in menselijke cellen21,22,23,24. Ten derde, SpCas9 is veruit het beste gekarakteriseerd enzym tot nu toe. Als een onderzoeker wil gebruiken een ander Cas nuclease, zou hij of zij vaak onduidelijk over de beste manier om het experiment en hoe goed de andere enzymen zal presteren in verschillende biologische contexten, in vergelijking met SpCas9 te ontwerpen.
Om duidelijkheid aan de relatieve prestaties van de verschillende systemen van de CRISPR-Cas, hebben we onlangs uitgevoerd een systematische vergelijking van vijf Cas enzym-SpCas9, het Cas9-enzym van Staphylococcus aureus (SaCas9), het enzym van de Cas9 van Neisseria meningitidis (NmCas9), het Cas12a-enzym van Acidaminococcus sp. BV3L6 (AsCas12a), en het enzym van de Cas12a van de Lachnospiraceae bacterie ND2006 (LbCas12a)25. We geëvalueerd voor een eerlijke vergelijking, de diverse Cas nucleasen met behulp van dezelfde set doel sites en andere experimentele omstandigheden. De studie ook afgebakend ontwerpparameters voor elk CRISPR-Cas-systeem, die als een nuttige referentie voor gebruikers van de technologie dienen zou. Hier, beter in staat stellen onderzoekers gebruik te maken van de CRISPR-Cas systeem, wij bieden een stapsgewijze protocol voor optimale genoom engineering met verschillende Cas9 en Cas12a enzymen (Zie Figuur 1). Het protocol omvat niet alleen experimentele details maar ook belangrijke ontwerpoverwegingen te maximaliseren van de kans op een succesvolle genoom engineering resultaat in zoogdiercellen.
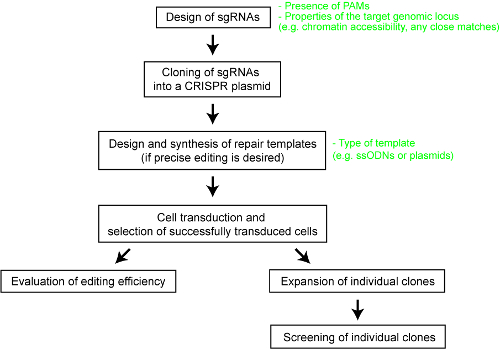
Figuur 1 : Een overzicht van de workflow voor het genereren van genoom bewerkt menselijke cellijnen. Klik hier voor een grotere versie van dit cijfer.
