An overview of the main steps for processing skin tissue to obtain a single cell suspension is shown in Figure 1, displaying the dermatoming of the skin (Figure 1A), different dermal layers (Figure 1B), removal of the subcutaneous fat layer (Figure 1C) and the separation of the epidermis and papillary dermis (Figure 1D), as well as the different steps of the manual and enzymatic tissue dissociation (Figure 1E,F). A scheme of the three dermal layers is provided in Figure 2.
Figure 3 displays the gating strategy of a flow cytometry staining panel for the analysis of different fibroblast subsets from human skin. Additional cell surface markers that are not expressed on fibroblasts permit the exclusion of various other cells present in the skin such as immune cells, epidermal cells, mesenchymal stem cells (MSCs), red blood cells or endothelial cells to attain maximal purity in the isolated populations. It is not critical to use the identical FACS panel used in Figure 2 for the identification of these fibroblast subpopulations, however this is our recommendation. One may use antibodies labelled with different fluorescent dyes or alter the combination of exclusion markers.
Of note, this protocol enables the identification of three fibroblast populations in human skin which present with different intradermal localization, gene expression profiles and also functions (Figure 4). FAP+CD90– are enriched in the papillary dermis whereas FAP+CD90+ and FAP–CD90+ are more abundant in the reticular dermis (Figure 4A). All three subpopulations exhibit the typical fibroblast morphology upon sorting in cell culture (Figure 4B). Interestingly, they differ regarding their ability to differentiate into adipocytes which is a hallmark for reticular fibroblasts. Combining these results with gene expression profiling via real-time polymerase chain reaction (RT-PCR)16 (Figure 4C), showing that FAP+CD90– cells express high levels of markers commonly attributed to papillary fibroblasts such as CD26, NTN and PDPN while CD90+ cells express the known reticular markers such as CD36, ACTA2 and PPARγ at high levels, we conclude that FAP+CD90– cells belong to the papillary and CD90+ cells belong to the reticular lineage.
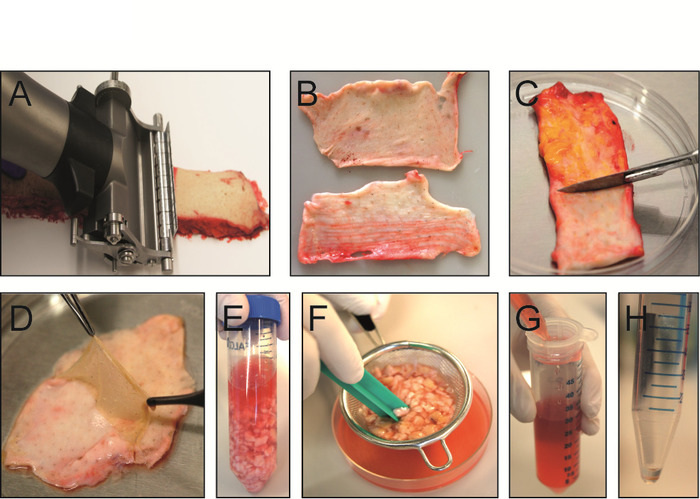
Figure 1: Isolation of dermal single cell suspension from intact human skin. (A) Skin is sliced with an electric dermatome into papillary and reticular dermis. (B) Papillary dermis with adjacent epidermis is 300 µm thick (top) while upper reticular dermis is 700 µm thick (bottom). (C) Subcutaneous fat layer is scraped-off of lower reticular dermis with a scalpel. (D) After incubation of papillary dermis in dissociation enzyme solution at 37 °C for 1 h, the epidermis can easily be removed with forceps. (E) Different dermal layers are minced with scissors and transferred into an enzyme digestion mix consisting of collagenase I, II and IV and hyaluronidase. (F) After 1 h of dissociation at 37 °C, tissue digestion is stopped and the suspension is poured through a tea strainer to remove undigested skin pieces. (G) Cell suspension is filtered once more through a 70 µm cell strainer and centrifuged at 500 x g at 4 °C for 10 min. (H) After centrifugation, the cell pellet is washed with 1x PBS with 10% FCS and is ready for FACS staining. Please click here to view a larger version of this figure.
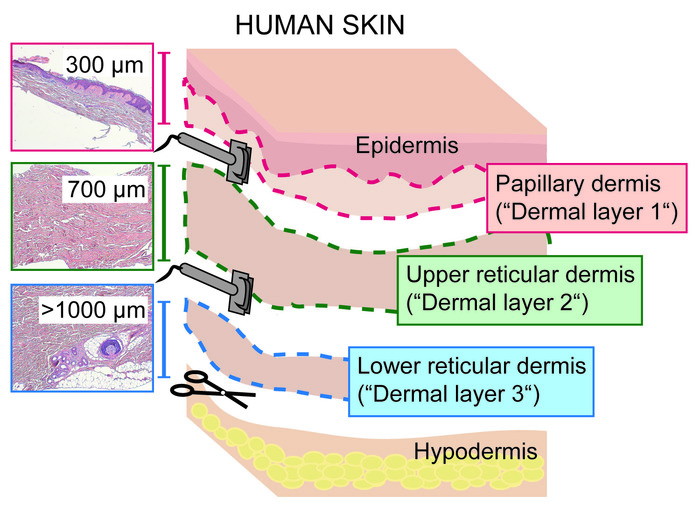
Figure 2: Sectioning of human skin dermis into papillary and reticular layers with a dermatome. Scheme showing the three dermal layers obtained by slicing full thickness skin into papillary dermis (including epidermis; 0‒300 µm; dermal layer 1), upper reticular (300‒1,000 µm; dermal layer 2) and lower reticular dermis (>1,000 µm; dermal layer 3) with a dermatome. Please click here to view a larger version of this figure.

Figure 3: FACS gating strategy for human dermal fibroblast subpopulations. (A‒E) First, cells are gated on single (B) and viable (DAPI–) cells (C). Immune cells (CD45+), mesenchymal stem cells (CD106+), red blood cells (CD235ab+) are excluded (C) and Pacific blue negative cells are gated further on E-cadherin (Ecad) and ITGA6 double negative cells (PE channel, D). E-cadherin and ITGA6 are markers expressed on epidermal cells. Next, CD31-FITC positive cells (endothelial and lymphatic cells) are excluded (D) resulting in three fibroblast populations expressing either one or both of the two cell surface fibroblast markers FAP and CD90: FAP+CD90–, FAP+CD90+ and FAP–CD90+ (E). Please click here to view a larger version of this figure.
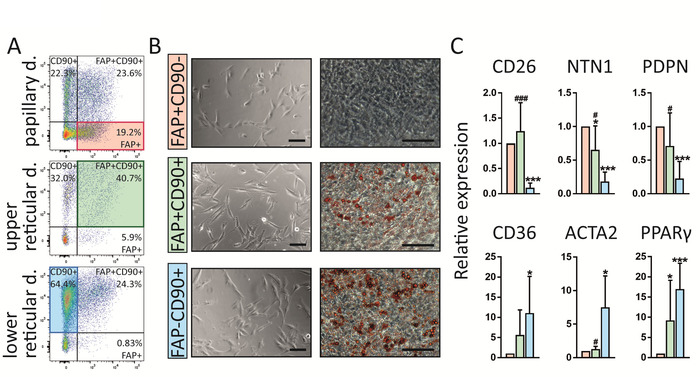
Figure 4: FAP+CD90–, FAP+CD90+ and FAP–CD90+ fibroblasts differ in dermal localization, gene expression and functionality. (A) Representative FACS plots of gated fibroblasts isolated from papillary, upper reticular and lower reticular dermis. Gating strategy is explained in Figure 3. FAP+CD90– fibroblasts are enriched in the papillary dermis (19.2%) compared to lower reticular dermis (0.83%). FAP+CD90+ cells can be found throughout the dermis but their highest abundance is in the upper reticular dermis (40.7%). FAP–CD90+ are enriched in the lower reticular dermis (64.4%). (B) Of note, all three sorted subpopulations display typical fibroblast morphology upon culture for 7 days (left). Interestingly, they behave differently in an adipogenesis assay. After 14 days in culture, FAP+CD90– do not differentiate into adipocytes while FAP+CD90+ and FAP–CD90+ readily undergo adipogenesis (right; Oil Red O stains lipid-bearing cells red). Scale bars = 1,000 µm. (C) Directly sorted FAP+CD90– fibroblasts express the papillary fibroblast markers CD26, NTN1 and PDPN, while FAP–CD90+ cells express CD36, ACTA2 and PPARγ, known to be expressed by the reticular lineage. * p ≤ 0.05; *** p ≤ 0.0005 compared to FAP+/CD90– cells; # p ≤ 0.05; ### p ≤ 0.0005 compared to FAP–/CD90+ cells. Please click here to view a larger version of this figure.
