Le gène de susceptibilité de type 1 du cancer du sein (BRCA1) est un gène suppresseur de tumeur largement connu. Puisque le gène BRCA1 est lié à la réparation des dommages d’ADN, les mutations dans ce gène conduirait à un plus grand risque de développement de cancer dans unindividu 1. Les cancers du sein, de l’ovaire, de la prostate et du pancréas sont liés à des mutations héréditaires de perte de fonction (LOF) du gène BRCA1 2. L’évaluation fonctionnelle et l’identification des variantes BRCA1 peuvent aider à prévenir et à diagnostiquer les diverses maladies. Pour répondre à la fonction des variantes BRCA1, plusieurs méthodes ont été développées et largement utilisées pour étudier la pathogénie des variantes BRCA1 telles que les essais de viabilité des cellules souches embryonnaires, les essais de reporter fluorescent et les essais thérapeutiques de sensibilité à base demédicaments 3,4,5,6. Bien que ces méthodes aient évalué la fonction d’un grand nombre de variantes BRCA1, les méthodes impliquant des variantes BRCA1 exprimées exogènement posent des limites en termes de surexpression qui pourraient affecter la régulation en aval, la posologie des gènes et le pliage desprotéines 7. En outre, ces analyses ne peuvent pas être exploitées au règlement posttranscriptionnel tel que l’épissage d’ARNm, la stabilité de transcription, et l’effet de la régionnon traduite 8,9.
Le système CRISPR-Cas9 permet l’édition ciblée du génome dans les cellules vivantes et lesorganismes 10. Grâce à un ARN à guide unique, Cas9 peut induire des ruptures à double brin (DSB) dans l’ADN chromosomique à des loci génomiques spécifiques afin d’activer deux voies de réparation de l’ADN : la voie de jointage final non homologue sujette aux erreurs (NHEJ) et la voie de réparation dirigée par homologie sans erreur (HDR)11. HDR est un mécanisme de réparation précis; cependant, les DSBs induits par la nucléase de Cas9 pour HDR ont souvent comme conséquence l’insertion et la mutation non désirées d’insertion et de suppression (indélébile). En outre, il a besoin de modèles d’ADN de donneur homologue pour réparer les dommages causés par l’ADN et a une efficacité relativement faible. Récemment, Cas9 nickase (nCas9) ont été fusionnés avec des domaines deaminase de cytidine pour cibler les substitutions C:G à T:A, sans avoir besoin de modèles d’ADN homologues et de ruptures de double brin d’ADN12,13,14,15. En utilisant l’éditeur de base de cytosine, nous avons développé une nouvelle méthode pour l’analyse fonctionnelle des variantes BRCA116.
Dans cette étude, nous avons utilisé l’éditeur de base de cytosine crispr-négocié, BE314, qui induit des mutations efficaces de C:G à T:A point, pour mettre en œuvre l’évaluation fonctionnelle des variantes BRCA1 et a identifié avec succès les fonctions de plusieurs variantes BRCA1 (Figure 1).
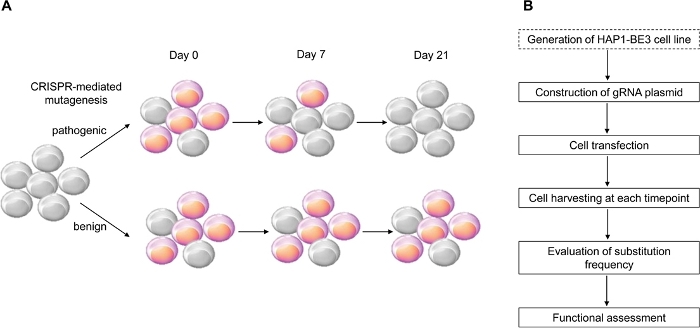
Figure 1 : Aperçu du flux de travail pour l’évaluation fonctionnelle. (A) Schéma montrant l’évaluation fonctionnelle de BRCA1. Puisque le LOF de BRCA1 affecte la viabilité cellulaire, quand la mutation BRCA1 est pathogène, les cellules meurent pendant que le nombre de passage augmente. (B) Étapes de l’évaluation fonctionnelle de BRCA1. La boîte pointillée est facultative. Il peut être remplacé par la co-transfection de l’expression de l’ARN g et be3 exprimant l’ADN des plasmides. S’il vous plaît cliquez ici pour voir une version plus grande de ce chiffre.
