Here we present two assays that measure C. albicans virulence as a non-lethal phenotype using C. elegans as an infection model. The first assay, fecundity, monitors how C. albicans infection impacts single hosts for progeny production and survival. The second assay, lineage expansion, measures how C. albicans infection impacts population growth over multiple generations.
The fecundity assay has multiple measures of host fitness during C. albicans infection. To assess how C. albicans infection impacts two distinct measures of host fitness, survival, and reproduction, we developed the fecundity assay19, which monitors individual hosts. Briefly, single hosts are isolated to control plates (uninfected) or C. albicans treatment plates and monitored every day for seven days for survival, and the number of daily progeny produced (Figure 1A). As this assay encompasses multiple measures of host fitness, it is an efficient way to assess C. albicans virulence. First, we evaluated total brood size by calculating the sum of daily progeny per host. Hosts infected with wildtype C. albicans produce a significantly smaller brood size on average compared to uninfected hosts (Figure 1B, black vs grey bars, p < 0.0001, Kuskal-Wallis test, and post-hoc Dunn's multiple comparison test). By monitoring daily progeny production, we can also detect differences in the timing of reproduction. We previously demonstrated that C. albicans infected hosts have delayed reproduction, with a large fraction of their progeny produced later in their adulthood compared to uninfected hosts19. The fraction of late reproduction is calculated by dividing the progeny produced on Days 4-7 by the total number of progeny produced. Uninfected hosts produce ~20% of their progeny during this late reproduction window whereas wildtype C. albicans infected hosts have 60% of their total offspring in this late reproduction window (Figure 1C).
The fecundity assay not only provides data to assess non-lethal virulence phenotypes, the data can also be used to assess host survival in a seven-day period. We plotted host survival (Figure 1D) and observed a decrease in survival for wildtype C. albicans infected hosts compared to uninfected hosts (grey vs. black lines, Figure 1D), but this difference was not statistically significant (p = 0.687, Log-rank test). This can be attributed to the short seven-day window of time this experiment covers. We previously showed it takes eight days for C. albicans infected hosts to reach 50% mortality19. Thus, this assay can only detect differences in host survival at very early time points.
Fecundity assays capture how C. albicans infection alters host fitness during early adulthood. We used this assay to screen for differences in virulence across reproductive and survival phenotypes using two C. albicans deletion strains (Table 1) previously identified to decrease virulence in murine, D. melanogaster, and C. elegans infection models11,19,23,24,25. CAS5 is a gene that encodes a transcription factor that regulates cell wall homeostasis, adherence, and stress response26. Here, hosts infected with C. albicans cas5Δ/Δ have significantly larger brood sizes and a smaller fraction of late reproduction compared to wildtype C. albicans infected hosts (Figure 1B,C, orange bars, p < 0.0001, Kruskal-Wallis test, and post-hoc Dunn's multiple comparison test). Additionally, hosts infected with C. albicanscas5Δ/Δ do not have significantly different brood sizes or fraction of late reproduction compared to uninfected hosts (p > 0.9999, Kruskal-Wallis test, and post-hoc Dunn's multiple comparison test), indicating that cas5Δ/Δ strains are avirulent. Furthermore, we found that cas5Δ/Δ reduces host mortality compared to wildtype C. albicans (Figure 1D, orange), although this difference is not statistically significant (p = 0.687, Log-rank test), likely due to the short timeframe in which host fitness is evaluated. RIM101 is a gene that encodes a transcription factor required for alkaline-induced hyphal growth27. Hosts infected with rim101Δ/Δ C. albicans had a significantly smaller fraction of late reproduction compared to wildtype C. albicans (Figure 1C, blue bar, p = 0.0184, Kruskal-Wallis test, and post-hoc Dunn's multiple comparison test) infected hosts, despite having similar total brood sizes (Figure 1B, blue bar, p = 0.6979, Kruskal-Wallis test, and post-hoc Dunn's multiple comparison test). Additionally, rim101Δ/Δ infected hosts had similar mortality to hosts infected with wildtype C. albicans (Figure 1D, blue, p = 0.687, Log-rank test). Taken together, we demonstrated the utility of this assay to distinguish subtle difference between C. albicans strains.
To quickly assess how C. albicans infection impacts host fecundity and survival over three generations, we developed the lineage expansion assay19, which monitors the progeny production of individual hosts. Briefly, single hosts are isolated to control plates (uninfected) or C. albicans treatment plates and after seven days the total number of viable progeny in the F1 and F2 generations are counted (Figure 2A). Uninfected hosts produced a progeny population ~35,000 in this timeframe (Figure 2B), compared to wildtype C. albicans infected hosts which produced a progeny population size of ~25,000, nearly a 30% reduction (Figure 2B). This simple assay can be used to rapidly screen through mutant strains of C. albicans. Both cas5Δ/Δ and rim101Δ/Δ C. albicans infected hosts produced progeny populations that were significantly larger than wildtype C. albicans infected hosts (Figure 2B, p < 0.0001 & p = 0.0185 respectively, Tukey's multiple comparisons test). Furthermore, cas5Δ/Δ C. albicans infected hosts produced progeny populations that were comparable to uninfected hosts, suggesting that this C. albicans strain is avirulent, and the rim101Δ/Δ C. albicans strain has reduced virulence for this virulence phenotype.
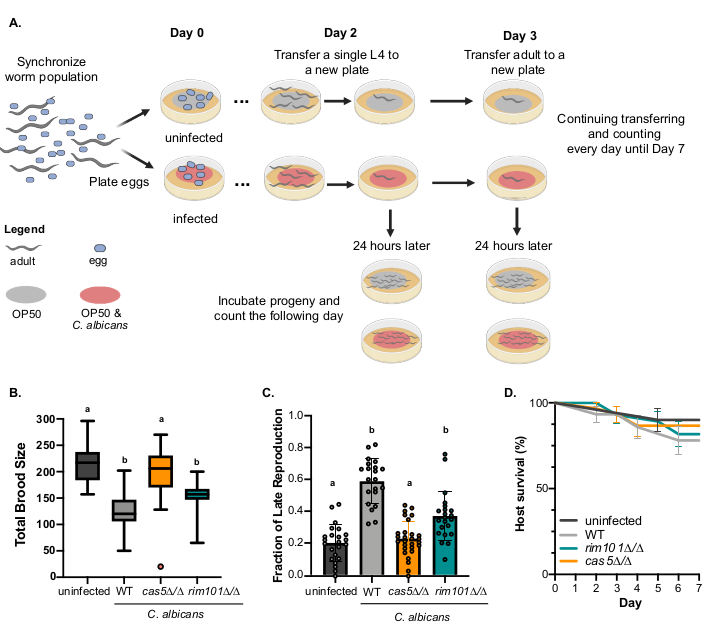
Figure 1: Fecundity and survival assessed in single hosts infected with different C. albicans strains. A) Experimental schematic of the fecundity assay. B) Total brood size for uninfected (OP50) hosts (n = 23), and hosts infected with wildtype (WT; SN250, n = 21), cas5Δ/Δ (n = 26), and rim101Δ/Δ (n = 26) C. albicans strains. The box represents the interquartile range, the midline indicates the median, and the whiskers represent the range. Error bars are the normalized range of the data. Treatments that share letters are not significantly differ, whereas treatments with differing letters are statistically significant, Kruskal-Wallis test and post-hoc Dunn's multiple comparison test. C) Fraction of late reproduction for uninfected hosts, and hosts infected with wildtype, cas5Δ/Δ, and rim101Δ/Δ C. albicans strains. Bars represent the mean, error bars represent +/- 1 SD, and symbols represent individuals hosts. Treatments that share letters are not statistically significantly different, whereas treatments with differing letters are significant, Kruskal-Wallis test and post-hoc Dunn's multiple comparison test. D) Survival curves of uninfected hosts, and hosts infected with wildtype, cas5Δ/Δ, and rim101Δ/Δ C. albicans strains for the first seven days of adulthood. Error bars represent ± 1 SD. Data from B, C, and D were collected from the same experiment and host sample sizes are the same in each panel. Please click here to view a larger version of this figure.
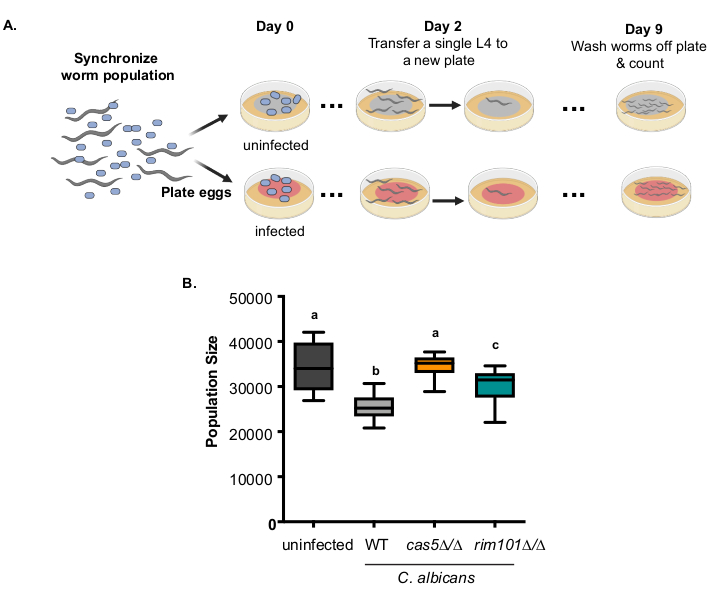
Figure 2: Virulence of C. albicans strains measured by host reproduction and death over multiple generations A) Experimental schematic of the lineage expansion assay. B) Box and whiskers plot of the population size (representing the number of F1 and F2 progeny) produced within 7 days from a single founder host exposed to OP50 food source alone (uninfected n=10, black), WT C. albicans (SN250, n=12, grey) or C. albicans cas5ΔΔ (n=12, orange), and rim101ΔΔ (n=12, teal) mutant strains (pink). Boxes indicate the 25-75th quartiles with median indicated. Error bars are the normalized range of the data. Treatments that share letters are not significantly different, whereas treatments with differing letters are statistically significant, one-way ANOVA and post-hoc Tukey multiple comparison test. Data was initially published in Feistel et al. 201916. Please click here to view a larger version of this figure.
| Strain | Genotype | Source |
| SN250 (WT) | his1Δ/his1Δ, leu2Δ::C.dubliniensis HIS1/leu2Δ::C.maltosa LEU2, arg4Δ /arg4Δ, URA3/ura3Δ::imm434, IRO1/iro1Δ::imm434 | Noble et al. (2010) |
| rim101∆/∆ | his1Δ/his1Δ, leu2Δ::C.dubliniensis HIS1/leu2Δ::C.maltosa LEU2, arg4Δ /arg4Δ, URA3/ura3Δ::imm434, IRO1/iro1Δ::imm434 orf19.7247Δ::C.dubliniensisHIS1/orf19.7247Δ::C.maltosaLEU2 | Noble et al. (2010) |
| cas5∆/∆ | his1Δ/his1Δ, leu2Δ::C.dubliniensis HIS1/leu2Δ::C.maltosa LEU2, arg4Δ /arg4Δ, URA3/ura3Δ::imm434, IRO1/iro1Δ::imm434 orf19.4670Δ::C.dubliniensisHIS1/orf19.4670Δ::C.maltosaLEU2 | Noble et al. (2010) |
Table 1: C. albicans strains used in this study
Supplemental Table 1: Sample Fecundity Data Please click here to download this table.
Supplemental Table 2: Sample Lineage Expansion Data Please click here to download this table.
