Las células inmunes se defienden contra patógenos y células cancerosas rastreando y escaneando continuamente las superficies de las células diana en busca de antígenos, tachonando su superficie 1,2. El reconocimiento de antígenos se inicia tras la unión entre el receptor de células T (TCR) y el complejo MHC del complejo de histocompatibilidad péptido-mayor (pMHC) expresado en la superficie de las células diana. Debido a que el reconocimiento TCR-pMHC ocurre en la unión entre dos células móviles, durante mucho tiempo se ha sospechado que experimenta fuerzas mecánicas. Además, esto condujo al modelo mecanosensor de activación TCR, que sugiere que las fuerzas TCR contribuyen a su función 3,4. Para comprender cuándo, dónde y cómo las fuerzas mecánicas contribuyen a la función de las células T, es imperativo desarrollar herramientas para visualizar las fuerzas moleculares transmitidas por las células T. Tradicionalmente, métodos como la microscopía de fuerza de tracción (TFM) y las matrices de micropilares se utilizan para investigar las fuerzas celulares 5,6. Sin embargo, la sensibilidad a la fuerza de las matrices TFM y micropilares está en la escala nanonewton (nN) y, por lo tanto, a menudo es insuficiente para estudiar las fuerzas moleculares de piconewton (pN) transmitidas por los receptores celulares7. Para mejorar la fuerza y la resolución espacial para la detección, nuestro laboratorio fue pionero en el desarrollo de sondas de tensión molecular, que inicialmente se sintetizaron utilizando polímeros de polietilenglicol (PEG)7. Las sondas de tensión molecular están compuestas por un “resorte” molecular extensible (PEG, proteína, ADN) flanqueado por un fluoróforo y un quencher y están anclados en una superficie. Las fuerzas aplicadas al extremo de la sonda conducen a su extensión, separando el fluoróforo y el exprimidor, generando así una fuerte señal de fluorescencia (Figura 1A)8,9,10.
Durante la última década hemos desarrollado una biblioteca de diferentes clases de sondas de tensión molecular con elementos de resorte hechos de ácidos nucleicos11, proteínas10 y polímeros8. Entre estos, las sondas de tensión basadas en ADN proporcionan la mayor relación señal / ruido y la mayor sensibilidad de fuerza, que se sintoniza fácilmente desde unos pocos pN hasta ~ 20 pN11. Hemos utilizado estas sondas de tensión de ADN en tiempo real para estudiar las fuerzas moleculares generadas por muchos tipos de células diversas, incluidos fibroblastos, células cancerosas, plaquetas y células inmunes11,12,13. Este manuscrito describirá protocolos para sintetizar y ensamblar sondas de tensión de ADN en una superficie para mapear las fuerzas del receptor molecular con resolución de fuerza pN utilizando un microscopio de fluorescencia convencional. Si bien el procedimiento actual incluye modificaciones químicas al ácido nucleico para introducir el reportero fluorescente (Figura 1B), es importante tener en cuenta que muchos de los pasos de modificación y purificación pueden subcontratarse a compañías personalizadas de síntesis de ADN. Por lo tanto, la tecnología de sondas de tensión de ADN es fácil y accesible para las comunidades más amplias de biología celular y mecanobiología.
Brevemente, para ensamblar sensores de tensión de ADN, una horquilla de ADN se hibrida con una hebra de ligando fluorescente en un brazo y una hebra de anclaje de quencher en el otro brazo y luego se inmoviliza en un sustrato de vidrio (Figura 1C, tensión en tiempo real). En ausencia de fuerza mecánica, la horquilla se cierra y, por lo tanto, la fluorescencia se apaga. Sin embargo, cuando la fuerza mecánica aplicada es mayor que F1/2 (la fuerza en equilibrio que conduce a una probabilidad del 50% de despliegue), la horquilla se derrite mecánicamente y se genera una señal fluorescente.
Sobre la base del sensor de tensión de ADN en tiempo real, también describimos protocolos para mapear las fuerzas acumuladas, lo que es particularmente útil para estudiar las interacciones entre los receptores en las células inmunes y su ligando natural. Esto se debe a que los receptores inmunes a menudo muestran enlaces de corta duración 3,14. Las fuerzas acumuladas se visualizan utilizando una hebra de “bloqueo” que se une preferentemente a horquillas de ADN abiertas y permite el almacenamiento de señales de fluorescencia asociadas con eventos de tracción mecánica (Figura 1C, tensión bloqueada). La hebra de bloqueo está diseñada para unir un sitio de unión críptico que se expone al derretir mecánicamente la horquilla inducida mecánicamente y bloquear la horquilla en estado abierto bloqueando el plegado de la horquilla, almacenando así la señal de tensión y generando un mapa de tensión acumulada. Además, la hebra de bloqueo está diseñada con un punto de apoyo de ocho nucleótidos, que permite una reacción de desplazamiento de la hebra mediada por la puntera con su complemento completo, la hebra de “desbloqueo”. Con la adición de la hebra de desbloqueo, la hebra de bloqueo atada se quita de la construcción de horquilla, borrando la señal de tensión almacenada y restableciendo la horquilla al estado en tiempo real.
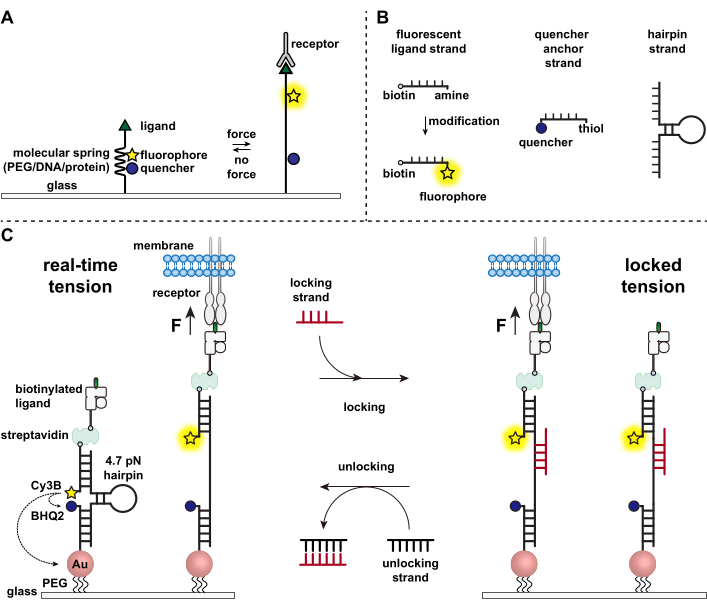
Figura 1: Esquema de las sondas de tensión molecular de última generación. (A) Diseño general de sondas de tensión molecular en tiempo real, (B) hebras para la construcción de la sonda de tensión basada en ADN, y (C) sondas de tensión basadas en ADN diseñadas y su alternancia entre el estado en tiempo real y el estado bloqueado. Haga clic aquí para ver una versión más grande de esta figura.
El protocolo principal consta de cuatro secciones principales: preparación de oligonucleótidos, preparación de superficies, imágenes y análisis de datos. Este protocolo ha sido demostrado con éxito por nuestro laboratorio y otros en células T OT-1 CD8 + ingenuas y activadas, células OT-II CD4 +, así como hibridomas, y se puede aplicar para interrogar diferentes receptores de células inmunes, incluidos el receptor de células T, el receptor de muerte celular programada (PD1) y las fuerzas del antígeno 1 asociado a la función de los linfocitos (LFA-1). Las células T OT-1 CD8 + no naïve se utilizan como una línea celular de ejemplo en este documento.
