تدافع الخلايا المناعية ضد مسببات الأمراض والخلايا السرطانية عن طريق الزحف المستمر ومسح أسطح الخلايا المستهدفة بحثا عن المستضدات ، وترصيع سطحها 1,2. يبدأ التعرف على المستضد عند الارتباط بين مستقبل الخلايا التائية (TCR) ومركب التوافق النسيجي الببتدي الرئيسي MHC (pMHC) المعبر عنه على سطح الخلايا المستهدفة. نظرا لأن التعرف على TCR-pMHC يحدث عند التقاطع بين خليتين متحركتين ، فقد اشتبه منذ فترة طويلة في أنه يعاني من قوى ميكانيكية. علاوة على ذلك ، أدى ذلك إلى نموذج المستشعر الميكانيكي لتنشيط TCR ، مما يشير إلى أن قوى TCR تساهم في وظيفتها 3,4. لفهم متى وأين وكيف تساهم القوى الميكانيكية في وظيفة الخلايا التائية ، من الضروري تطوير أدوات لتصور القوى الجزيئية التي تنتقل بواسطة الخلايا التائية. تقليديا ، يتم استخدام طرق مثل مجهر قوة الجر (TFM) ومصفوفات micropillar للتحقيق في القوى الخلوية 5,6. ومع ذلك ، فإن حساسية القوة لمصفوفات TFM و micropillar تكون على مقياس nanonewton (nN) ، وبالتالي فهي غالبا ما تكون غير كافية لدراسة قوى piconewton الجزيئية (pN) التي تنتقل بواسطة مستقبلات الخلايا7. لتحسين القوة والدقة المكانية للكشف ، كان مختبرنا رائدا في تطوير مجسات التوتر الجزيئي ، والتي تم تصنيعها في البداية باستخدام بوليمرات البولي إيثيلين جلايكول (PEG)7. تتكون مجسات التوتر الجزيئي من “زنبرك” جزيئي قابل للتمديد (PEG ، بروتين ، حمض نووي) محاط بفلوروفور ومروي ومثبت على سطح. تؤدي القوى المطبقة على نهاية المسبار إلى امتداده ، وفصل الفلوروفور والتبريد ، وبالتالي توليد إشارة مضان قوية (الشكل 1 أ)8،9،10.
على مدار العقد الماضي ، قمنا بتطوير مكتبة من فئات مختلفة من مجسات التوتر الجزيئي مع عناصر زنبركية مصنوعة من الأحماض النووية11 والبروتينات10 والبوليمرات8. من بين هذه ، توفر مجسات التوتر القائمة على الحمض النووي أعلى نسبة إشارة إلى الضوضاء وأكبر حساسية للقوة ، والتي يمكن ضبطها بسهولة من بضعة pN إلى ~ 20 pN11. لقد استخدمنا مجسات توتر الحمض النووي في الوقت الفعلي لدراسة القوى الجزيئية الناتجة عن العديد من أنواع الخلايا المتنوعة ، بما في ذلك الخلايا الليفية والخلايا السرطانية والصفائح الدموية والخلايا المناعية11،12،13. ستصف هذه المخطوطة بروتوكولات لتجميع وتجميع مجسات توتر الحمض النووي على سطح لرسم خريطة لقوى المستقبلات الجزيئية بدقة قوة pN باستخدام مجهر مضان تقليدي. في حين أن الإجراء الحالي يتضمن تعديلات كيميائية على الحمض النووي لإدخال مراسل الفلورسنت (الشكل 1 ب) ، من المهم ملاحظة أن العديد من خطوات التعديل والتنقية يمكن الاستعانة بمصادر خارجية لشركات تخليق الحمض النووي المخصصة. لذلك ، فإن تقنية مجسات توتر الحمض النووي سهلة ، ويمكن الوصول إليها من قبل مجتمعات بيولوجيا الخلية والبيولوجيا الميكانيكية الأوسع.
باختصار ، لتجميع مستشعرات توتر الحمض النووي ، يتم تهجين دبوس شعر الحمض النووي إلى حبلا يجند فلوري على ذراع واحدة وشريط مرساة مروي على الذراع الأخرى ثم يجمد على ركيزة زجاجية (الشكل 1 ج ، التوتر في الوقت الفعلي). في حالة عدم وجود قوة ميكانيكية ، يتم إغلاق دبوس الشعر ، وبالتالي يتم إخماد التألق. ومع ذلك ، عندما تكون القوة الميكانيكية المطبقة أكبر من F1/2 (القوة عند التوازن التي تؤدي إلى احتمال 50٪ للتكشف) ، يذوب دبوس الشعر ميكانيكيا ، ويتم إنشاء إشارة فلورسنت.
بناء على مستشعر توتر الحمض النووي في الوقت الفعلي، نصف أيضا بروتوكولات لرسم خريطة للقوى المتراكمة، وهو أمر مفيد بشكل خاص لدراسة التفاعلات بين المستقبلات على الخلايا المناعية ورباطها الطبيعي. وذلك لأن المستقبلات المناعية غالبا ما تعرض روابط قصيرة العمر 3,14. يتم تصوير القوى المتراكمة باستخدام حبلا “قفل” يرتبط بشكل تفضيلي بفتح دبابيس شعر الحمض النووي ويسمح بتخزين إشارات التألق المرتبطة بأحداث السحب الميكانيكية (الشكل 1C ، التوتر المقفل). تم تصميم حبلا القفل لربط موقع ربط خفي يتعرض عند ذوبان دبوس الشعر المستحث ميكانيكيا وقفل دبوس الشعر في الحالة المفتوحة عن طريق منع إعادة طي دبوس الشعر ، وبالتالي تخزين إشارة التوتر ، وتوليد خريطة توتر متراكمة. علاوة على ذلك ، تم تصميم حبلا القفل بإصبع قدم مكون من ثمانية نيوكليوتيدات ، مما يتيح تفاعل إزاحة حبلا بوساطة إصبع القدم مع مكمله الكامل ، حبلا “فتح”. مع إضافة حبلا الفتح ، يتم تجريد حبلا القفل المربوط من بنية دبوس الشعر ، مما يؤدي إلى محو إشارة التوتر المخزنة وإعادة ضبط دبوس الشعر إلى حالة الوقت الفعلي.
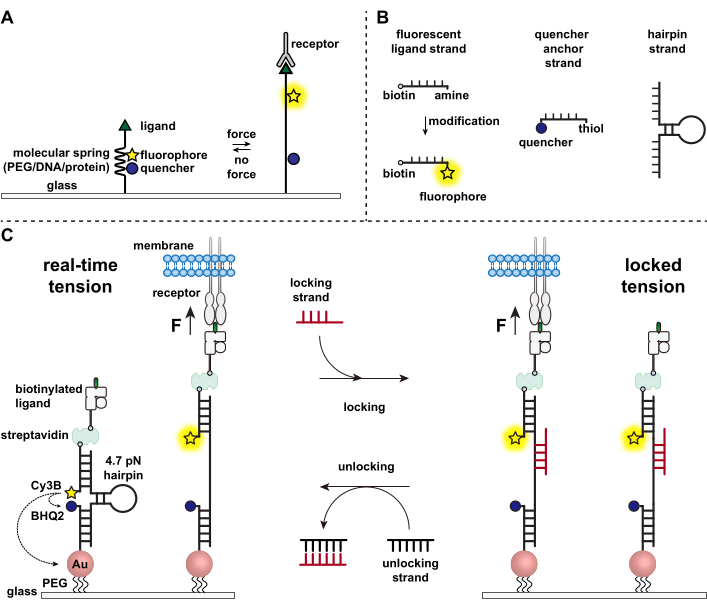
الشكل 1: مخطط أحدث مجسات التوتر الجزيئي. (أ) التصميم العام لمسبار التوتر الجزيئي في الوقت الفعلي، (ب) خيوط بناء مسبار التوتر القائم على الحمض النووي، (ج) مجسات التوتر القائمة على الحمض النووي (DNA) وتبديلها بين الحالة في الوقت الفعلي والحالة المقفلة. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
يتكون البروتوكول الرئيسي من أربعة أقسام رئيسية – إعداد قليل النوكليوتيد ، وإعداد السطح ، والتصوير ، وتحليل البيانات. تم إثبات هذا البروتوكول بنجاح من قبل مختبرنا وغيره في خلايا OT-1 CD8 + T الساذجة والمنشطة ، وخلايا OT-II CD4 + ، وكذلك الأورام الهجينة ، ويمكن تطبيقه لاستجواب مستقبلات الخلايا المناعية المختلفة بما في ذلك مستقبلات الخلايا التائية ، ومستقبلات موت الخلايا المبرمجة (PD1) ، وقوى المستضد 1 المرتبطة بوظيفة الخلايا الليمفاوية (LFA-1). تستخدم الخلايا التائية الساذجة OT-1 CD8 + كمثال على خط الخلية في هذه الورقة.
