Immunceller forsvarer sig mod patogener og kræftceller ved kontinuerligt at kravle og scanne overfladerne af målceller for antigener og studse deres overflade 1,2. Antigengenkendelse initieres ved binding mellem T-cellereceptoren (TCR) og peptid-major histokompatibilitetskomplekset MHC (pMHC) kompleks udtrykt på overfladen af målceller. Fordi TCR-pMHC-genkendelse sker ved krydset mellem to mobile celler, har det længe været mistænkt for at opleve mekaniske kræfter. Desuden førte dette til mekanosensormodellen for TCR-aktivering, hvilket tyder på, at TCR-kræfter bidrager til dens funktion 3,4. For at forstå, hvornår, hvor og hvordan mekaniske kræfter bidrager til T-cellefunktionen, er det bydende nødvendigt at udvikle værktøjer til at visualisere de molekylære kræfter, der transmitteres af T-celler. Traditionelt bruges metoder som traction force mikroskopi (TFM) og micropillar arrays til at undersøge cellulære kræfter 5,6. Imidlertid er kraftfølsomheden af TFM og mikrosøjle arrays på nanonewton (nN) skalaen og er derfor ofte utilstrækkelig til at studere de molekylære piconewton (pN) kræfter, der transmitteres af cellereceptorer7. For at forbedre kraften og den rumlige opløsning til detektion var vores laboratorium banebrydende for udviklingen af molekylære spændingsprober, som oprindeligt blev syntetiseret ved hjælp af polyethylenglycol (PEG) polymerer7. Molekylspændingssonder består af en udvidelig molekylær “fjeder” (PEG, protein, DNA) flankeret af en fluorofor og slukker og er forankret på en overflade. Kræfter, der påføres sondens endestation, fører til dens forlængelse, adskiller fluoroforen og slukkeren og genererer således et stærkt fluorescenssignal (figur 1A)8,9,10.
I løbet af det sidste årti har vi udviklet et bibliotek af forskellige klasser af molekylære spændingsprober med fjederelementer fremstillet af nukleinsyrer11, proteiner10 og polymerer8. Blandt disse giver de DNA-baserede spændingssonder det højeste signal / støjforhold og den største kraftfølsomhed, som let indstilles fra nogle få pN op til ~ 20 pN11. Vi har brugt disse DNA-spændingssonder i realtid til at studere de molekylære kræfter, der genereres af mange forskellige celletyper, herunder fibroblaster, kræftceller, blodplader og immunceller11,12,13. Dette manuskript vil beskrive protokoller til syntetisering og samling af DNA-spændingssonder på en overflade for at kortlægge molekylære receptorkræfter med pN-kraftopløsning ved hjælp af et konventionelt fluorescensmikroskop. Mens den nuværende procedure inkluderer kemiske modifikationer af nukleinsyren for at introducere den fluorescerende reporter (figur 1B), er det vigtigt at bemærke, at mange af modifikations- og oprensningstrinnene kan outsources til brugerdefinerede DNA-syntesevirksomheder. Derfor er DNA-spændingssondeteknologi let og tilgængelig for de bredere cellebiologi- og mekanobiologiske samfund.
Kort sagt, for at samle DNA-spændingssensorer, hybridiseres en DNA-hårnål til en fluorescerende ligandstreng på den ene arm og en slukningsankerstreng på den anden arm og immobiliseres derefter på et glassubstrat (figur 1C, realtidsspænding). I mangel af mekanisk kraft lukkes hårnålen, og fluorescensen slukkes således. Men når den påførte mekaniske kraft er større end F1/2 (kraften ved ligevægt, der fører til en 50% sandsynlighed for udfoldelse), smelter hårnålen mekanisk, og der genereres et fluorescerende signal.
Med udgangspunkt i DNA-spændingssensoren i realtid beskriver vi også protokoller til kortlægning af akkumulerede kræfter, hvilket er særligt nyttigt til at studere interaktioner mellem receptorer på immunceller og deres naturlige ligand. Dette skyldes, at immunreceptorer ofte viser kortvarige bindinger 3,14. Akkumulerede kræfter afbildes ved hjælp af en “låsende” streng, der fortrinsvis binder til åbne DNA-hårnåle og muliggør lagring af fluorescenssignaler forbundet med mekaniske trækhændelser (figur 1C, låst spænding). Låsestrengen er designet til at binde et kryptisk bindingssted, der eksponeres ved mekanisk induceret smeltning af hårnålen og låse hårnålen i åben tilstand ved at blokere hårnålens foldning igen, således at spændingssignalet lagres og genererer et akkumuleret spændingskort. Desuden er låsestrengen designet med et otte-nukleotid-tåhold, som muliggør en tåholdsmedieret strengforskydningsreaktion med sit fulde komplement, “oplåsningsstrengen”. Med tilføjelsen af oplåsningsstrengen fjernes den bundne låsestreng fra hårnålekonstruktionen, hvilket sletter det lagrede spændingssignal og nulstiller hårnålen tilbage til realtidstilstanden.
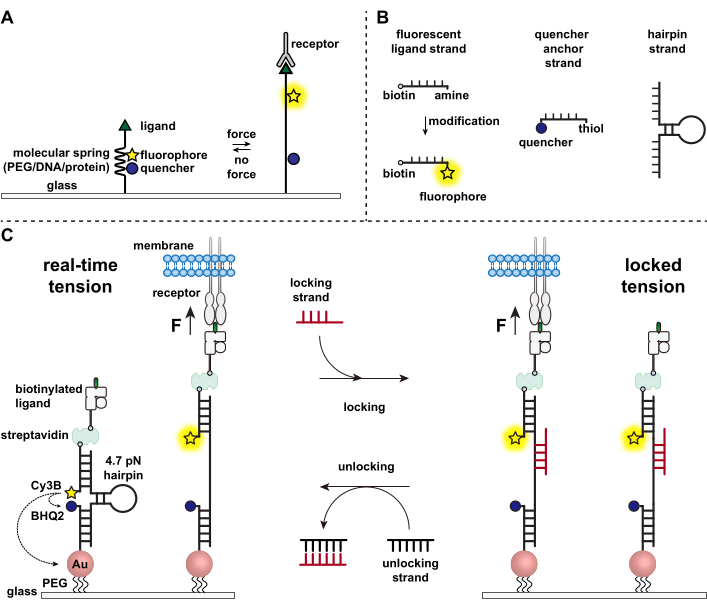
Figur 1: Skema for de nyeste molekylære spændingssonder. (A) Generelt design af molekylær spændingssonde i realtid, (B) strenge til den DNA-baserede spændingssondekonstruktion og (C) konstruerede DNA-baserede spændingssonder og deres skift mellem realtidstilstand og låst tilstand. Klik her for at se en større version af denne figur.
Hovedprotokollen består af fire hovedafsnit – oligonukleotidpræparation, overfladebehandling, billeddannelse og dataanalyse. Denne protokol er med succes blevet demonstreret af vores laboratorium og andre i naive og aktiverede OT-1 CD8 + T-celler, OT-II CD4 + celler samt hybridomer og kan anvendes til at forhøre forskellige immuncellereceptorer, herunder T-cellereceptor, programmeret celledødsreceptor (PD1) og lymfocytfunktionsassocieret antigen 1 (LFA-1) kræfter. OT-1 CD8+ naive T-celler bruges som eksempel på cellelinje i dette papir.
