Les cellules immunitaires se défendent contre les agents pathogènes et les cellules cancéreuses en rampant et en balayant continuellement les surfaces des cellules cibles à la recherche d’antigènes, en cloutant leur surface 1,2. La reconnaissance de l’antigène est initiée lors de la liaison entre le récepteur des lymphocytes T (TCR) et le complexe peptide-majeur d’histocompatibilité CMH (pMHC) exprimé à la surface des cellules cibles. Parce que la reconnaissance TCR-pMHC se produit à la jonction entre deux cellules mobiles, il a longtemps été soupçonné de subir des forces mécaniques. De plus, cela a conduit au modèle mécanocapteur d’activation TCR, qui suggère que les forces TCR contribuent à sa fonction 3,4. Pour comprendre quand, où et comment les forces mécaniques contribuent au fonctionnement des lymphocytes T, il est impératif de développer des outils pour visualiser les forces moléculaires transmises par les lymphocytes T. Traditionnellement, des méthodes telles que la microscopie à force de traction (TFM) et les réseaux de micropiliers sont utilisés pour étudier les forces cellulaires 5,6. Cependant, la sensibilité aux forces de la TFM et des réseaux de micropiliers est à l’échelle du nanonewton (nN) et est donc souvent insuffisante pour étudier les forces moléculaires du piconewton (pN) transmises par les récepteurs cellulaires7. Pour améliorer la force et la résolution spatiale de la détection, notre laboratoire a été le pionnier du développement de sondes de tension moléculaire, qui ont été initialement synthétisées à l’aide de polymères de polyéthylèneglycol (PEG)7. Les sondes de tension moléculaire sont composées d’un « ressort » moléculaire extensible (PEG, protéine, ADN) flanqué d’un fluorophore et d’un quencher et sont ancrées sur une surface. Les forces appliquées à l’extrémité de la sonde conduisent à son extension, séparant le fluorophore et l’anti-quenceur, et générant ainsi un fort signal de fluorescence (Figure 1A)8,9,10.
Au cours de la dernière décennie, nous avons développé une bibliothèque de différentes classes de sondes de tension moléculaire avec des éléments à ressort fabriqués à partir d’acides nucléiques11, de protéines10 et de polymères8. Parmi celles-ci, les sondes de tension basées sur l’ADN fournissent le rapport signal sur bruit le plus élevé et la plus grande sensibilité à la force, qui est facilement réglée de quelques pN à ~20 pN11. Nous avons utilisé ces sondes de tension d’ADN en temps réel pour étudier les forces moléculaires générées par de nombreux types de cellules, y compris les fibroblastes, les cellules cancéreuses, les plaquettes et les cellules immunitaires11,12,13. Ce manuscrit décrira des protocoles pour synthétiser et assembler des sondes de tension d’ADN sur une surface afin de cartographier les forces des récepteurs moléculaires avec une résolution de force pN à l’aide d’un microscope à fluorescence conventionnel. Bien que la procédure actuelle comprenne des modifications chimiques de l’acide nucléique pour introduire le rapporteur fluorescent (figure 1B), il est important de noter que de nombreuses étapes de modification et de purification peuvent être sous-traitées à des entreprises de synthèse d’ADN personnalisées. Par conséquent, la technologie des sondes de tension de l’ADN est facile et accessible aux communautés plus larges de la biologie cellulaire et de la mécanobiologie.
Brièvement, pour assembler des capteurs de tension d’ADN, une épingle à cheveux à ADN est hybridée à un brin de ligand fluorescent sur un bras et à un brin d’ancrage de quencher sur l’autre bras, puis immobilisée sur un substrat de verre (Figure 1C, tension en temps réel). En l’absence de force mécanique, l’épingle à cheveux est fermée et la fluorescence est donc éteinte. Cependant, lorsque la force mécanique appliquée est supérieure à la F1/2 (la force à l’équilibre qui conduit à une probabilité de 50% de se déplier), l’épingle à cheveux fond mécaniquement et un signal fluorescent est généré.
En nous appuyant sur le capteur de tension de l’ADN en temps réel, nous décrivons également des protocoles pour cartographier les forces accumulées, ce qui est particulièrement utile pour étudier les interactions entre les récepteurs des cellules immunitaires et leur ligand naturel. En effet, les récepteurs immunitaires présentent souvent des liaisons de courte durée 3,14. Les forces accumulées sont imagées à l’aide d’un brin de « verrouillage » qui se lie préférentiellement aux épingles à cheveux d’ADN ouvertes et permet le stockage des signaux de fluorescence associés aux événements de traction mécanique (Figure 1C, tension verrouillée). Le brin de verrouillage est conçu pour lier un site de liaison cryptique qui est exposé lors de la fusion induite mécaniquement de l’épingle à cheveux et verrouiller l’épingle à cheveux à l’état ouvert en bloquant le repliement de l’épingle à cheveux, stockant ainsi le signal de tension et générant une carte de tension accumulée. De plus, le brin de verrouillage est conçu avec un toehold à huit nucléotides, ce qui permet une réaction de déplacement du brin médiée par le teil avec son complément complet, le brin de « déverrouillage ». Avec l’ajout du brin de déverrouillage, le brin de verrouillage lié est retiré de la construction en épingle à cheveux, effaçant le signal de tension stocké et réinitialisant l’épingle à cheveux à l’état temps réel.
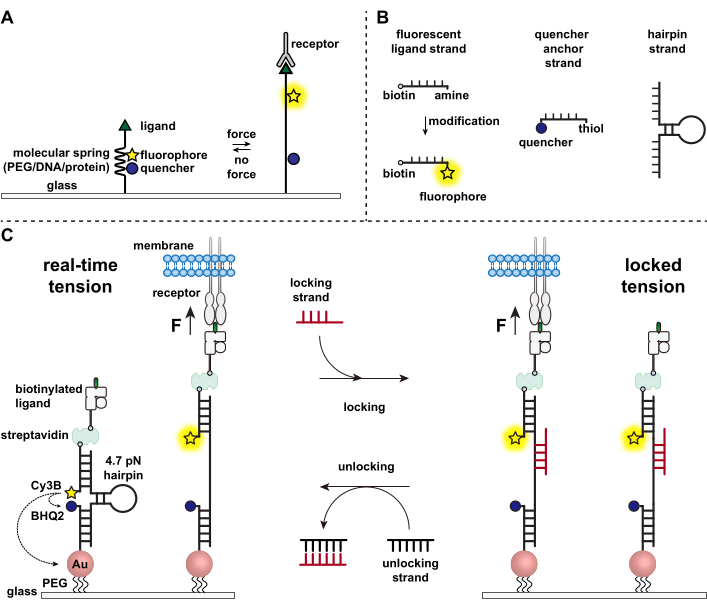
Figure 1 : Schéma des sondes de tension moléculaire de pointe. (A) Conception générale de sonde de tension moléculaire en temps réel, (B) Brins pour la construction de sonde de tension basée sur l’ADN, et (C) sondes de tension basées sur l’ADN et leur basculement entre l’état en temps réel et l’état verrouillé. Veuillez cliquer ici pour voir une version agrandie de cette figure.
Le protocole principal comprend quatre sections principales: préparation des oligonucléotides, préparation de surface, imagerie et analyse des données. Ce protocole a été démontré avec succès par notre laboratoire et d’autres dans des lymphocytes T CD8+ OT-1 naïfs et activés, des cellules CD4+ OT-II, ainsi que des hybridomes, et peut être appliqué pour interroger différents récepteurs de cellules immunitaires, y compris le récepteur des cellules T, le récepteur de mort cellulaire programmée (PD1) et les forces de l’antigène 1 associé à la fonction lymphocytaire (LFA-1). Les lymphocytes T naïfs de CD8+ OT-1 sont utilisés comme exemple de lignée cellulaire dans cet article.
