Immunceller forsvarer seg mot patogener og kreftceller ved kontinuerlig å krype og skanne overflatene til målceller for antigener, og pigge overflaten 1,2. Antigengjenkjenning initieres ved binding mellom T-cellereseptoren (TCR) og peptid-major histokompatibilitetskomplekset MHC (pMHC) komplekset uttrykt på overflaten av målceller. Fordi TCR-pMHC-gjenkjenning skjer i krysset mellom to mobilceller, har det lenge vært mistanke om å oppleve mekaniske krefter. Videre førte dette til mekanosensormodellen for TCR-aktivering, noe som antyder at TCR-krefter bidrar til funksjonen 3,4. For å forstå når, hvor og hvordan mekaniske krefter bidrar til T-cellefunksjon, er det viktig å utvikle verktøy for å visualisere molekylære krefter som overføres av T-celler. Tradisjonelt brukes metoder som trekkraftmikroskopi (TFM) og mikropillararrays for å undersøke cellulære krefter 5,6. Imidlertid er kraftfølsomheten til TFM og mikropilarrayer på nanonewton (nN) -skalaen og er derfor ofte utilstrekkelig til å studere molekylære piconewton (pN) -krefter overført av cellereseptorer7. For å forbedre kraften og romlig oppløsning for deteksjon, var laboratoriet vårt banebrytende for utviklingen av molekylære spenningsprober, som opprinnelig ble syntetisert ved bruk av polyetylenglykol (PEG) polymerer7. Molekylære spenningsprober består av en utvidbar molekylær “fjær” (PEG, protein, DNA) flankert av en fluorofor og slukker og er forankret på en overflate. Krefter påført sondens endepunkt fører til dens forlengelse, separerer fluoroforen og slukkeren, og genererer dermed et sterkt fluorescenssignal (figur 1A) 8,9,10.
I løpet av det siste tiåret har vi utviklet et bibliotek med forskjellige klasser av molekylære spenningsprober med fjærelementer laget av nukleinsyrer11, proteiner10 og polymerer8. Blant disse gir de DNA-baserte spenningsprobene det høyeste signal-støyforholdet og den største kraftfølsomheten, som enkelt justeres fra noen få pN opp til ~20 pN11. Vi har brukt disse sanntids DNA-spenningsprobene til å studere molekylære krefter generert av mange forskjellige celletyper, inkludert fibroblaster, kreftceller, blodplater og immunceller11,12,13. Dette manuskriptet vil beskrive protokoller for å syntetisere og sette sammen DNA-spenningsprober på en overflate for å kartlegge molekylære reseptorkrefter med pN-kraftoppløsning ved hjelp av et konvensjonelt fluorescensmikroskop. Mens den nåværende prosedyren inkluderer kjemiske modifikasjoner av nukleinsyren for å introdusere fluorescerende reporter (figur 1B), er det viktig å merke seg at mange av modifikasjons- og rensetrinnene kan outsources til tilpassede DNA-syntesefirmaer. Derfor er DNA-spenningssondeteknologien lett og tilgjengelig for de bredere cellebiologi- og mekanobiologimiljøene.
Kort sagt, for å montere DNA-spenningssensorer, hybridiseres en DNA-hårnål til en fluorescerende ligandstreng på den ene armen og en slukkeankerstreng på den andre armen og immobiliseres deretter på et glasssubstrat (figur 1C, sanntidsspenning). I fravær av mekanisk kraft er hårnålen lukket, og dermed slokkes fluorescensen. Men når den påførte mekaniske kraften er større enn F1/2 (kraften ved likevekt som fører til en 50% sannsynlighet for å utfolde seg), smelter hårnålen mekanisk, og et fluorescerende signal genereres.
Ved å bygge på sanntids DNA-spenningssensor beskriver vi også protokoller for å kartlegge akkumulerte krefter, noe som er spesielt nyttig for å studere interaksjoner mellom reseptorer på immunceller og deres naturlige ligand. Dette skyldes at immunreseptorer ofte viser kortvarige bindinger 3,14. Akkumulerte krefter avbildes ved hjelp av en “låsende” streng som fortrinnsvis binder seg til åpne DNA-hårnåler og muliggjør lagring av fluorescenssignaler assosiert med mekaniske trekkhendelser (figur 1C, låst spenning). Låsestrengen er designet for å binde et kryptisk bindingssted som eksponeres ved mekanisk indusert smelting av hårnålen og låse hårnålen i åpen tilstand ved å blokkere hårnålsfolding, og dermed lagre spenningssignalet og generere et akkumulert spenningskart. Videre er låsestrengen designet med et åtte-nukleotidtåhold, som muliggjør en tåholdsmediert strengforskyvningsreaksjon med sitt fulle komplement, den “låsende” strengen. Med tillegg av opplåsingsstrengen fjernes den bundne låsestrengen av hårnålskonstruksjonen, sletter det lagrede spenningssignalet og tilbakestiller hårnålen tilbake til sanntidstilstanden.
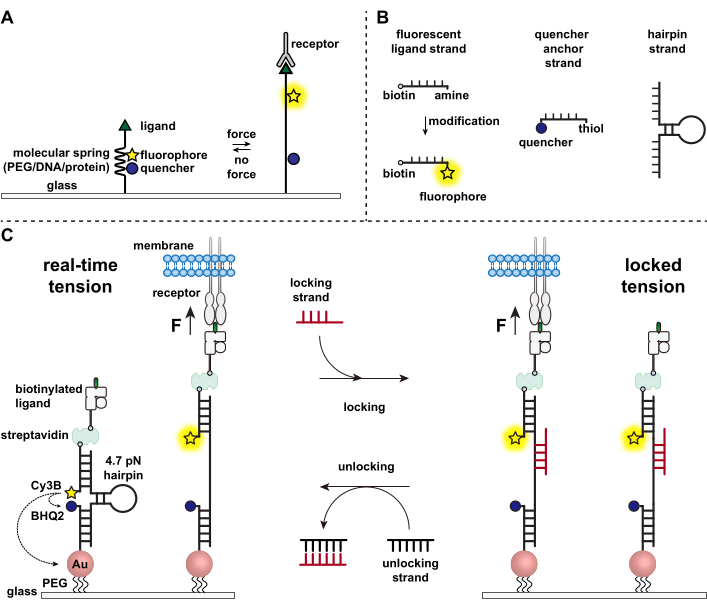
Figur 1: Skjema for state-of-art molekylære spenningsprober. (A) Generell utforming av sanntids molekylær spenningssonde, (B) Tråder for DNA-basert spenningssondekonstruksjon, og (C) konstruerte DNA-baserte spenningsprober og deres veksling mellom sanntidstilstand og låst tilstand. Klikk her for å se en større versjon av denne figuren.
Hovedprotokollen består av fire hovedseksjoner – oligonukleotidpreparasjon, overflatebehandling, avbildning og dataanalyse. Denne protokollen har blitt vellykket demonstrert av laboratoriet vårt og andre i naive og aktiverte OT-1 CD8 + T-celler, OT-II CD4 + -celler, samt hybridomer, og kan brukes til å forhøre forskjellige immuncellereseptorer, inkludert T-cellereseptor, programmert celledødsreseptor (PD1) og lymfocyttfunksjonsassosiert antigen 1 (LFA-1) krefter. OT-1 CD8+ naive T-celler brukes som eksempelcellelinje i denne artikkelen.
