Иммунные клетки защищаются от патогенов и раковых клеток, непрерывно ползая и сканируя поверхности клеток-мишеней на наличие антигенов, усеивая их поверхность 1,2. Распознавание антигена инициируется при связывании между Т-клеточным рецептором (TCR) и комплексом пептидно-главной гистосовместимости MHC (pMHC), экспрессируемым на поверхности клеток-мишеней. Поскольку распознавание TCR-pMHC происходит на стыке между двумя подвижными клетками, долгое время подозревалось, что он испытывает механические силы. Более того, это привело к механосенсорной модели активации TCR, которая предполагает, что силы TCR способствуют его функции 3,4. Чтобы понять, когда, где и как механические силы способствуют функционированию Т-клеток, необходимо разработать инструменты для визуализации молекулярных сил, передаваемых Т-клетками. Традиционно для исследования клеточных сил5,6 используются такие методы, как тракционно-силовая микроскопия (TFM) и микропиллярные матрицы. Однако силовая чувствительность TFM и микростолбовых матриц находится в масштабе наноньютона (nN) и, таким образом, часто недостаточна для изучения молекулярных пиконьютонных (pN) сил, передаваемых клеточными рецепторами7. Чтобы улучшить силу и пространственное разрешение для обнаружения, наша лаборатория впервые разработала зонды молекулярного натяжения, которые первоначально были синтезированы с использованием полимеров полиэтиленгликоля (ПЭГ)7. Зонды молекулярного натяжения состоят из выдвижной молекулярной «пружины» (ПЭГ, белок, ДНК), окруженной флуорофором и гасителем, и закреплены на поверхности. Силы, приложенные к концу зонда, приводят к его расширению, разделяя флуорофор и гаситель, и, таким образом, генерируя сильный сигнал флуоресценции (рис. 1А)8,9,10.
За последнее десятилетие мы разработали библиотеку различных классов зондов молекулярного натяжения с пружинными элементами, изготовленными из нуклеиновых кислот11, белков10 и полимеров8. Среди них датчики натяжения на основе ДНК обеспечивают самое высокое соотношение сигнал/шум и наибольшую чувствительность к силе, которая легко настраивается от нескольких пН до ~20 пН11. Мы использовали эти зонды натяжения ДНК в реальном времени для изучения молекулярных сил, генерируемых многими различными типами клеток, включая фибробласты, раковые клетки, тромбоциты и иммунные клетки11,12,13. В этой рукописи будут описаны протоколы синтеза и сборки зондов натяжения ДНК на поверхности для картирования молекулярных рецепторных сил с разрешением силы pN с использованием обычного флуоресцентного микроскопа. В то время как текущая процедура включает химические модификации нуклеиновой кислоты для введения флуоресцентного репортера (рис. 1B), важно отметить, что многие этапы модификации и очистки могут быть переданы на аутсорсинг компаниям, занимающимся синтезом ДНК. Таким образом, технология зондов натяжения ДНК проста и доступна для более широких сообществ клеточной биологии и механобиологии.
Вкратце, для сборки датчиков натяжения ДНК шпилька ДНК гибридизуется с флуоресцентной цепью лиганда на одном плече и якорной нитью гасителя на другом плече, а затем иммобилизуется на стеклянной подложке (рис. 1C, натяжение в реальном времени). При отсутствии механической силы шпилька закрывается, и таким образом флуоресценция гасится. Однако, когда приложенная механическая сила больше, чем F1/2 (сила в равновесии, которая приводит к 50% вероятности разворачивания), шпилька механически плавится, и генерируется флуоресцентный сигнал.
Основываясь на датчике натяжения ДНК в реальном времени, мы также описываем протоколы для отображения накопленных сил, что особенно полезно для изучения взаимодействия между рецепторами на иммунных клетках и их естественным лигандом. Это связано с тем, что иммунные рецепторы часто демонстрируют короткоживущие связи 3,14. Накопленные силы визуализируются с помощью «запирающей» цепи, которая преимущественно связывается с открытыми шпильками ДНК и позволяет хранить флуоресцентные сигналы, связанные с механическими событиями вытягивания (рис. 1C, заблокированное натяжение). Фиксирующая прядь предназначена для связывания загадочного места связывания, который подвергается воздействию при механически индуцированном плавлении шпильки, и блокировки шпильки в открытом состоянии путем блокировки повторного складывания шпильки, тем самым сохраняя сигнал натяжения и генерируя накопленную карту натяжения. Кроме того, блокирующая цепь имеет восьминуклеотидную опору, которая обеспечивает реакцию смещения цепи, опосредованную опорой на носок, с ее полным дополнением, «разблокирующей» нитью. С добавлением отпирающей пряди связанная фиксирующая прядь снимается с конструкции шпильки, стирая сохраненный сигнал натяжения и сбрасывая шпильку обратно в состояние реального времени.
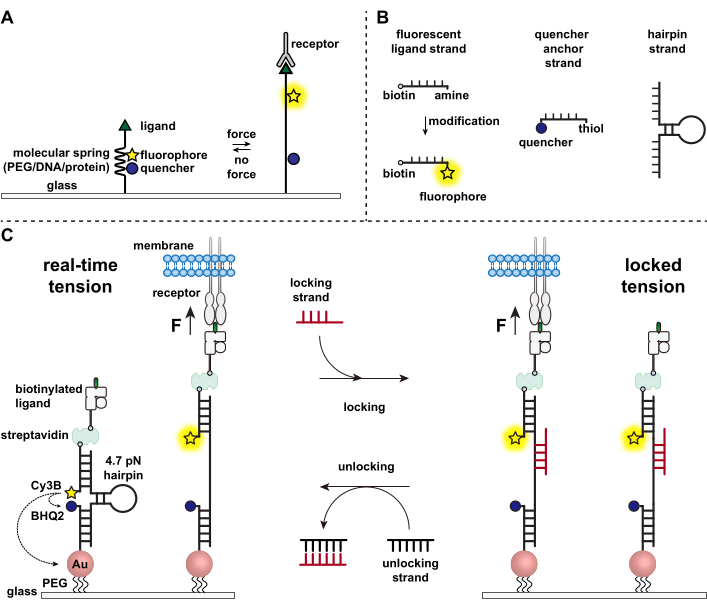
Рисунок 1: Схема современных датчиков молекулярного натяжения. (A) Общая конструкция датчика молекулярного натяжения в реальном времени, (B) Нити для конструкции датчика натяжения на основе ДНК и (C) спроектированные датчики натяжения на основе ДНК и их переключение между состоянием реального времени и заблокированным состоянием. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Основной протокол состоит из четырех основных разделов – подготовка олигонуклеотидов, подготовка поверхности, визуализация и анализ данных. Этот протокол был успешно продемонстрирован нашей лабораторией и другими в наивных и активированных OT-1 CD8+ Т-клетках, OT-II CD4+ клетках, а также гибридомах и может применяться для опроса различных рецепторов иммунных клеток, включая рецептор Т-клеток, рецептор запрограммированной клеточной смерти (PD1) и силы антигена 1, связанного с функцией лимфоцитов (LFA-1). OT-1 CD8+ наивные Т-клетки используются в качестве примера клеточной линии в этой статье.
