Immunceller försvarar sig mot patogener och cancerceller genom att kontinuerligt krypa och skanna målcellernas ytor för antigener, studding deras yta 1,2. Antigenigenkänning initieras vid bindning mellan T-cellreceptorn (TCR) och det peptid-stora histokompatibilitetskomplexet MHC (pMHC) -komplexet uttryckt på ytan av målceller. Eftersom TCR-pMHC-igenkänning sker vid korsningen mellan två mobila celler har den länge misstänkts för att uppleva mekaniska krafter. Dessutom ledde detta till mekanosensormodellen för TCR-aktivering, vilket tyder på att TCR-krafter bidrar till dess funktion 3,4. För att förstå när, var och hur mekaniska krafter bidrar till T-cellernas funktion är det absolut nödvändigt att utveckla verktyg för att visualisera de molekylära krafter som överförs av T-celler. Traditionellt används metoder som dragkraftmikroskopi (TFM) och mikropelarrayer för att undersöka cellulära krafter 5,6. Emellertid är kraftkänsligheten hos TFM- och mikropelarrayer på nanonewton (nN) -skalan och är därför ofta otillräcklig för att studera molekylära piconewton (pN) -krafter som överförs av cellreceptorer7. För att förbättra kraften och den rumsliga upplösningen för detektion var vårt laboratorium banbrytande för utvecklingen av molekylära spänningsprober, som ursprungligen syntetiserades med användning av polyetylenglykol (PEG) polymerer7. Molekylära spänningsprober består av en utdragbar molekylär “fjäder” (PEG, protein, DNA) flankerad av en fluorofor och släckare och förankras på en yta. Krafter som appliceras på sondens ändpunkt leder till dess förlängning, separerar fluoroforen och släckaren och genererar därmed en stark fluorescenssignal (figur 1A)8,9,10.
Under det senaste decenniet har vi utvecklat ett bibliotek av olika klasser av molekylära spänningsprober med fjäderelement tillverkade av nukleinsyror11, proteiner10 och polymerer8. Bland dessa ger de DNA-baserade spänningsproberna det högsta signal-brusförhållandet och den största kraftkänsligheten, som enkelt ställs in från några pN upp till ~ 20 pN11. Vi har använt dessa DNA-spänningsprober i realtid för att studera de molekylära krafterna som genereras av många olika celltyper, inklusive fibroblaster, cancerceller, blodplättar och immunceller11,12,13. Detta manuskript kommer att beskriva protokoll för att syntetisera och montera DNA-spänningsprober på en yta för att kartlägga molekylära receptorkrafter med pN-kraftupplösning med hjälp av ett konventionellt fluorescensmikroskop. Medan det nuvarande förfarandet inkluderar kemiska modifieringar av nukleinsyran för att introducera den fluorescerande reportern (figur 1B), är det viktigt att notera att många av modifierings- och reningsstegen kan outsourcas till anpassade DNA-syntesföretag. Därför är DNA-spänningsprober teknik lätt och tillgänglig för de bredare cellbiologi- och mekanobiologiska samhällena.
Kortfattat, för att montera DNA-spänningssensorer, hybridiseras en DNA-hårnål till en fluorescerande ligandsträng på ena armen och en släckarankarsträng på den andra armen och immobiliseras sedan på ett glassubstrat (Figur 1C, realtidsspänning). I frånvaro av mekanisk kraft stängs hårnålen och fluorescensen släckes. Men när den applicerade mekaniska kraften är större än F1/2 (kraften vid jämvikt som leder till en 50% sannolikhet för utfällning) smälter hårnålen mekaniskt och en fluorescerande signal genereras.
Med utgångspunkt i DNA-spänningssensorn i realtid beskriver vi också protokoll för att kartlägga ackumulerade krafter, vilket är särskilt användbart för att studera interaktioner mellan receptorer på immunceller och deras naturliga ligand. Detta beror på att immunreceptorer ofta uppvisar kortlivade bindningar 3,14. Ackumulerade krafter avbildas med hjälp av en “låsande” sträng som företrädesvis binder till öppna DNA-hårnålar och möjliggör lagring av fluorescenssignaler associerade med mekaniska draghändelser (figur 1C, låst spänning). Låssträngen är utformad för att binda ett kryptiskt bindningsställe som exponeras vid mekaniskt inducerad smältning av hårnålen och låsa hårnålen i öppet tillstånd genom att blockera hårnålens omvikning, vilket lagrar spänningssignalen och genererar en ackumulerad spänningskarta. Dessutom är låssträngen utformad med en åtta-nukleotid-tåhållare, vilket möjliggör en tåhållmedierad strängförskjutningsreaktion med sitt fulla komplement, den “upplåsta” strängen. Med tillägget av upplåsningssträngen avlägsnas den bundna låssträngen från hårnålskonstruktionen, raderar den lagrade spänningssignalen och återställer hårnålen till realtidsläget.
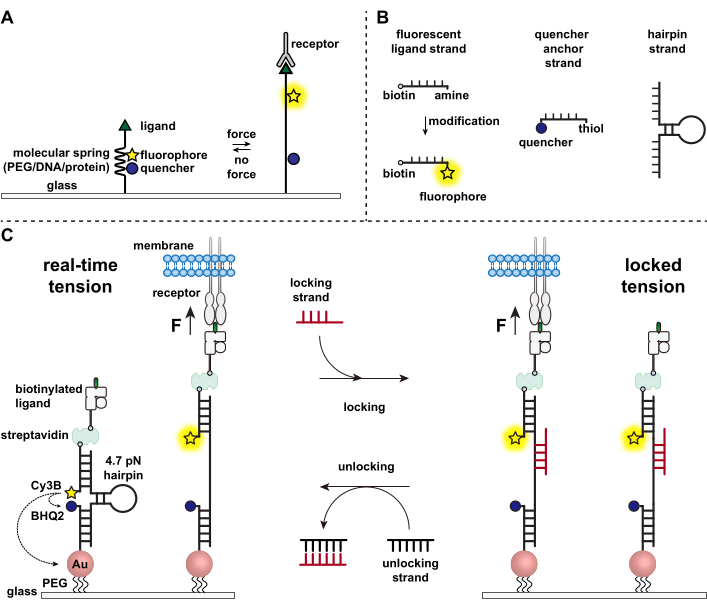
Figur 1: Schema för de toppmoderna molekylära spänningsproberna . (A) Allmän design av molekylär spänningssond i realtid, (B) strängar för den DNA-baserade spänningssondkonstruktionen, och (C) konstruerade DNA-baserade spänningssonder och deras växling mellan realtidstillstånd och låst tillstånd. Klicka här för att se en större version av denna figur.
Huvudprotokollet består av fyra huvudsektioner – oligonukleotidberedning, ytbehandling, avbildning och dataanalys. Detta protokoll har framgångsrikt demonstrerats av vårt laboratorium och andra i naiva och aktiverade OT-1 CD8 + T-celler, OT-II CD4 + -celler, såväl som hybridom, och kan tillämpas för att förhöra olika immuncellreceptorer inklusive T-cellreceptor, programmerad celldödsreceptor (PD1) och lymfocytfunktionsassocierad antigen 1 (LFA-1) krafter. OT-1 CD8 + naiva T-celler används som exempel cellinje i detta papper.
