Um grande obstáculo no estudo de doenças fibrosas é a falta de modelos representativos de tecido 3D humano que forneçam insights sobre o comportamento dos fibroblastos e seus derivados patológicos. Para estudar processos fibrosos, os sistemas de cultura 2D padrão são sub-ideais, uma vez que os fibroblastos isolados transdiferem rapidamente em α-smooth muscle actin (SMA) expressando miofibroblasts quando cultivados em substratos 2D não compatíveis1,2,3. Assim, os fibroblastos na cultura 2D padrão não refletem um fenótipo de tecido “saudável” regular3,4,5,6. Culturas em substratos flexíveis foram introduzidas para simular ambientes de tecidos não fibrosos (10 kPa) e fibrosos (35 kPa), mas estes não têm a terceira dimensão, o que é muito importante no que diz respeito à fisiopatologia. A engenharia de tecidos proporciona a oportunidade de superar essa limitação, permitindo a cultura do fibroblasto em um contexto de matriz extracelular (ECM) definida e experimentalmente, por alterações na celularidade, composição de ECM e concentração de ECM, tudo isso pode determinar a biomecânica tecidual.
Vários modelos 3D foram gerados usando fibroblastos. Discos flutuantes e microesferas estiveram entre os primeiros e demonstram que o colágeno é remodelado e compactado de forma dependente do tempo. Os fibroblastos exercem forças de tração em fibrilas de colágeno, um processo que pode ser facilitado pela adição de agentes pró-fibrosos, como a transformação do fator de crescimento-beta 1 (TGF-β1)8,9,10,11,12,13,14,15,16. No entanto, culturas livremente flutuantes não permitem o carregamento externo controlado e, portanto, constituem modelos de encolhimento ou compactação continuamente. Tecidos projetados em forma de folha abriram a possibilidade de estudar a regulação homeostática de propriedades biomecânicas dos tecidos, ou seja, através de testes de cepas uni, bi, multiaxial ou cíclica17,18,19,20. Esses modelos têm sido usados, por exemplo, para demonstrar a influência do número celular na rigidez tecidual, que foi encontrada para se correlacionar positivamente com a integridade do citoesqueleto e a contratilidade do citosqueleto18,19. No entanto, é importante notar que as conversões de força-a-tensão são complicadas pela deformação de tecido não uniforme em torno de pontos de fixação de transdutores de força e pontos de ancoragem. Essa limitação inerente pode ser contornada por tecidos em forma de osso de cachorro ou anel, oferecendo alguma aplicação tecidual em pontos de ancoragem21,22,23. Tecidos em forma de anel podem ser preparados distribuindo um hidrogel de colágeno celular em moldes em forma de anel. À medida que o hidrogel compacta, um tecido se forma em torno da haste interna descompressível do molde, que oferece resistência para maior contração tecidual24,25,26,27. Após a compactação inicial e tipicamente máxima, os tecidos também podem ser transferidos para espaçadores ajustáveis para conter ainda mais a ECT circular em um comprimento de tecido definido3,24,25,26,27,28,29,30. As propriedades biofísicas podem ser avaliadas em dispositivos de tensão horizontal ou vertical padrão com células de carga apropriadas sob tensão unidirecional ou dinâmica3. Como os tecidos possuem uma estrutura circular em grande parte uniforme e podem ser mantidos em barras/ganchos (pontos de ancoragem e/ou transdutores de força), embora estes ainda possam incluir áreas de compressão ao redor das barras de carregamento, este formato permite uma variação de tensão mais uniforme em comparação com a fixação3. Além disso, tecidos ancorados provocam uma forma celular bipolar, e as células se adaptam às forças teciduais pelo alongamento ao longo das linhas de força promovendo tração anisotrópico31,32,33,34,35,36. Nós já aplicamos ECT em forma de anel de fibroblastos cardíacos de ratos e humanos (CF) em torno de um único polo rígido em experimentos funcionais de tensão de estresse e realizamos ganhos e perdas de estudos de função usando fibroblastos viralmente transduzidos24,25,26 e estudos farmacológicos37. Além disso, poderíamos identificar diferenças sexuais na fibrose mediada por CF no modelo ECT27.
O seguinte protocolo para a geração de ECT humano, exemplificado com CF humano primário obtido como CF criopreservado de fornecedores comerciais (ver Tabela de Materiais), combina as vantagens dos tecidos em forma de anel com uma maneira fácil e rápida de produzir tecidos macroscópicos para uma plataforma de 48 poços projetada para testes paralelos de alto teor de conteúdo.
É importante ressaltar que o modelo ECT não se restringe a um tipo específico de fibroblasto, com o uso documentado na investigação de outros fibroblastos, por exemplo, fibroblastos de pele38,39. Além disso, os fibroblastos das biópsias do paciente funcionam igualmente bem, e a escolha dos fibroblastos depende, em última análise, da questão científica a ser abordada.
A plataforma utilizada para a geração de ECT descrita neste protocolo é uma placa de cultura celular/tecido 3D disponível comercialmente (Figura 1A). Os métodos para a formação, cultivo e monitoramento da formação e função ect sob uma geometria definida e carga mecânica com a ajuda da placa de 48 poços são descritos. A ECT formada é mantida por polos flexíveis integrados e a carga mecânica pode ser ajustada de acordo com o propósito final usando postes com dureza diferentes (Valor 36-89 da Costa A), influenciando suas rigidezs de dobra. Polos com uma costa Um valor de 46 são recomendados. O protocolo é, além disso, compatível com um molde circular personalizado descrito anteriormente, onde o ECT é mantido em torno de uma única haste rígida37. As dimensões deste molde são dadas na Figura 1B.
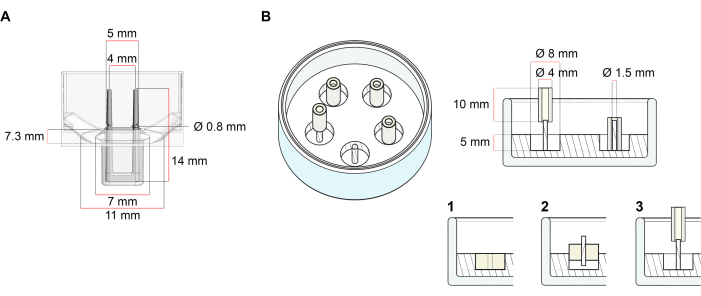
Figura 1: Representação esquemática dos moldes de fundição. (A) Desenho técnico e dimensões de um molde de fundição com dois polos flexíveis. O molde compreende uma circunferência interna delimitada por uma parede curta que contém polos de retenção duplas no corpo principal do molde. Os polos flexíveis têm uma distância horizontal livre entre si e estão conectados na base. O molde permite um volume de fundição de 180 μL. O poço de cada molde permite uma capacidade de volume de pelo menos 600 μL de mídia cultural. Diferentes composições materiais podem ser usadas para produzir polos com rigidezs específicas (por exemplo, TM5MED-TM9MED). (B) Desenho técnico e dimensões de um molde em forma de anel com uma única haste dura. Trata-se de um molde alternativo com geometria distinta e ambiente mecânico, que pode ser usado com o protocolo de fundição ECT37. O método de montagem do molde em forma de anel foi adaptado a partir de formatos maiores publicados28,41. Em suma, o método inclui (1) fixação de espaçadores de moldagem politetrafluoroetileno (PTFE) (8 mm de diâmetro) em polidimtilsiloxano (PDMS, silicone) derramado em pratos de vidro (diâmetro de 60 mm) e (2) fixação de um suporte de poste PDMS (1,5 mm de diâmetro) concentricamente dentro da cavidade oca formada, que serve para (3) segurar um poste removível (tubo de silicone de 4 mm de diâmetro). O resultante do espaço oco permite 180 μL de volume de fundição. Cada prato de vidro pode portar múltiplos moldes impressos (exemplarmente mostrados com 5 moldes) e tem capacidade para até 5 mL de meio de cultura. Clique aqui para ver uma versão maior desta figura.
