مثيلة الحمض النووي هي علامة جينية مهمة تنظم التعبير الجيني وبنية الكروماتين. يحدث المثيلة في الغالب في ثنائي النوكليوتيدات CpG – السيتوزين يليه جوانوزين. تضاف مجموعة الميثيل إلى موضع 5 من السيتوزين. هناك حاجة إلى أنماط مثيلة الحمض النووي الصحيحة ، وبالتالي التعبير الجيني المناسب ، من أجل التطور والوظيفة الخلوية المناسبة. ارتبطت العديد من حالات المرض بالتغيرات في نمط المثيلة الطبيعي1،2،3. على سبيل المثال ، هناك صلة بين بدء السرطان وتقدمه والتعديلات في نمط مثيلة الحمض النووي. عادة ، تظهر الخلايا السرطانية مستويات إجمالية أقل من ميثيل سيتوزين ، مما يساهم في عدم استقرار الجينوم. في الوقت نفسه ، يتركز ميثيل السيتوزين الموجود في الجينوم في المناطق المروجة للجينات المثبطة للورم ، مما يؤدي إلى إسكات الجينات لهذه البروتينات المهمة. والجدير بالذكر أن التغيرات اللاجينية ديناميكية وقابلة للعكس ، على عكس طفرات الحمض النووي المرتبطة بتكوين الأورام. وقد جعل هذا البروتينات المشاركة في تنظيم الجينات اللاجينية أهدافا دوائية مثيرة للاهتمام 2,4.
ميثيل ترانسفيراز الحمض النووي (DNMTs) هي البروتينات المسؤولة عن توليد والحفاظ على أنماط مثيلة الحمض النووي. توجد ثلاثة إيزوزيمات نشطة تحفيزيا ، DNMT1 و DNMT3a و DNMT3b ، في البشر. أثناء التطوير والتمايز ، تنشئ دي نوفو ميثيل ترانسفيراز ، DNMT3a و DNMT3b ، أنماط مثيلة. يمكن لكلا الإنزيمين ربط بروتين DNMT3L غير النشط تحفيزيا لتشكيل مجمعات تظهر نشاطا متزايدا 1,5. بعد انقسام الخلايا ، تحتوي الخلايا البنوية على الحمض النووي نصف الميثيل – الحمض النووي الذي يحتوي على ميثيل السيتوزين في شريط واحد فقط من الازدواج – لأن الحمض النووي المركب حديثا يخلو من علامات المثيلة. تتمثل الوظيفة الرئيسية ل DNMT1 في ميثيل هذا الحمض النووي نصف الميثيل ، وبالتالي إعادة إنشاء نمط المثيلة الكامل 1,5.
الروابط بين نشاط DNMT والسرطان راسخة. الإفراط في التعبير عن DNMT1 ، إما عن طريق آليات النسخ أو ما بعد الترجمة ، هو نتيجة للعديد من مسارات الأورام الشائعة6،7،8،9. تؤدي الأساليب الجينية لخفض نشاط DNMT1 باستخدام الأليلات ناقصة الشكل إلى انخفاض تكوين الورم في الفئران Apc (Min) 10. oligonucleotides المضادة للحساسية التي تمنع DNMT1 الأورام في زراعة الخلايا ونماذج ورم الفأر11,12. وبالتالي ، فإن تثبيط نشاط DNMT1 يبدو وكأنه نهج واعد لعلاج السرطان. ومع ذلك ، فإن الأدوار التي تلعبها إيزوزيمات DNMT3 ليست بهذه البساطة. تم العثور على طفرات DNMT3a في سرطان الدم النخاعي الحاد13 ومتلازمة خلل التنسج النقوي14. ثبت أن طفرة واحدة على الأقل من الطفرات المحددة تقلل من نشاط مثيلة الحمض النووي للإنزيم15. ومع ذلك ، يتم التعبير عن DNMT3b بشكل مفرط في سرطان الثدي16 وسرطان القولون والمستقيم17. مع لعب إيزوزيمات DNMT المختلفة أدوارا مختلفة في التسرطن ، فإن تحديد مثبطات الإيزوزيم الخاصة سيكون أمرا بالغ الأهمية. لن تكون هذه المركبات مفيدة فقط لتطوير العلاجات ، ولكن مثبطات الإيزوزيم الخاصة ستكون أيضا أداة لا تقدر بثمن لتشريح دور كل إيزوزيم DNMT في مسببات السرطان.
تم الإبلاغ عن العديد من مثبطات DNMT في الأدبيات. يمكن تقسيم مثبطات DNMT المعروفة إلى فئتين: النوكليوزيد وغير النيوكليوسيد. مثبطات النيوكليوزيد هي عادة نظائر السيتيدين. يتم دمج هذه المركبات في الحمض النووي وتحبس DNMTs تساهميا. تمت الموافقة على 5-azacytidine و 5-aza-2′-deoxycytidine لعلاج متلازمة خلل التنسج النقوي وسرطان الدم النخاعي الحاد4،18. تمثل السمية العالية والتوافر البيولوجي المنخفض وعدم الاستقرار الكيميائي لهذه المركبات مشاكل. العمل الجاري هو دراسة فعالية الجيل القادم من مثبطات النيوكليوسيد. SGI-110 ، المشتق من 5-aza-2′-deoxycytidine ، هو أحد الأمثلة19,20. مثبطات النيوكليوزيد ليست خاصة بالإيزوزيم وستعطل أي إيزوزيم DNMT تمت مواجهته. لذلك ، يؤدي العلاج بعامل إزالة الميثيل من النوكليوزيد إلى استنفاد جميع إيزوزيمات DNMT 4,18. لا يلزم دمج مثبطات غير النوكليوزيد في الحمض النووي لممارسة آثارها المثبطة. بدلا من ذلك ، ترتبط هذه الجزيئات مباشرة ب DNMTs ، مما يوفر إمكانية تثبيط خاص بالإيزوزيم. تم اكتشاف العديد من مثبطات غير النوكليوزيد حتى الآن ، بما في ذلك SGI-1027 21 ، و hydralazine 22 ، و procainamide 23 ، و RG108 ومشتقاته24 ، والمنتجات الطبيعية ، (-) -epigallocatechin 3-gallate (EGCG) 25 وحمض اللاكايكA 26,27. معظم مثبطات غير النوكليوزيد المكتشفة حتى الآن ليست انتقائية للأيزوزيم أو تظهر تفضيلات ضعيفة لإيزوزيم DNMT واحد. بالإضافة إلى ذلك ، يجب تحسين فاعلية هذه الجزيئات ، خاصة في الخلايا 4,18. وبالتالي ، هناك حاجة لاكتشاف أو تطوير مثبطات DNMT أكثر قوة وانتقائية للأيزوزيم.
تتمثل إحدى العقبات التي تحول دون اكتشاف مثبطات جزيئات صغيرة جديدة ل DNMTs في المقايسات الشاقة المستخدمة تقليديا لفحص نشاط DNMT28. عادة ما تكون المقايسات متقطعة بخطوات متعددة. لا يزال النشاط الأنزيمي ل DNMTs يتم فحصه بشكل روتيني باستخدام ميثيونين S-adenosyl المشع (SAM) 29،30،31،32،33،34. كما تم تطوير مقايسات غير مشعة لمثيلة الحمض النووي. على سبيل المثال ، تم وصف المقايسات التي تستخدم نوكلياز تقييد حساسية للميثيل والرحلان الكهربائي لفصل منتجات الهضم35,36. هذه الأنواع من المقايسات المتقطعة متعددة الخطوات ليست قابلة بسهولة لاكتشاف الأدوية. منذ منتصف عام 2000 ، تم تطوير العديد من فحوصات مثيلة الحمض النووي ذات الإنتاجية الأعلى28. تم استخدام اختبار القرب التلألؤ للكشف عن مثبطات DNMT137. تم استخدام اختبار آخر يستخدم نوكلياز تقييد حساسية للميثيل للكشف عن مثبطات DNMT3a25,38. في حين أن كلا الاختبارين يسمحان بإنتاجية أعلى من مقايسات مثيلة الحمض النووي التقليدية ، فإن المقايسات تتطلب خطوات متعددة ولا تسمح بمراقبة نشاط المثيلة في الوقت الفعلي. في الآونة الأخيرة ، تم وصف مقايسة حركية مستمرة تجمع بين تكوين S-adenosylhomocysteine (SAH) ، وهو أحد منتجات تفاعل المثيلة ، والتغير الطيفي عند 340 نانومتر المرتبط بأكسدة NADPH39. يستخدم هذا الفحص ثلاثة إنزيمات اقتران لتوليد إشارة طيفية.
لقد طورنا مقايسة مثيلة الحمض النووي المقترنة بنوكلياز داخلية قائمة على التألق والتي تستخدم إنزيم اقتران واحد متاح تجاريا ويمكنها توليد البيانات في الوقت الفعلي (الشكل 1). يستخدم قليل النوكليوتيد الذي يحتوي على ثلاثة ميثيل سيتوزين كركيزة. يحتوي الحمض النووي للركيزة على فلوروفور في الطرف 5 ‘ومطفئ في الطرف 3’. تولد مثيلة موقع CpG نصف الميثيل موقع الانقسام للنوكلياز الداخلي Gla I – GCGC الميثيلي بالكامل. Gla I انشقاق المنتج قليل النوكليوتيد يطلق الفلوروفور من المروي ويولد مضان في الوقت الحقيقي. يمكن استخدام الفحص لفحص نشاط أي شكل متساوي من DNMT ؛ ومع ذلك ، لوحظ نشاط أعلى مع DNMT1 حيث أن هذا الإيزوزيم يثيل بشكل تفضيلي الحمض النووينصف الميثيل 1,5. ويلاحظ نشاط أكثر قوة إذا تمت إزالة مجال تسلسل استهداف بؤر النسخ المتماثل (RFTS) المثبط ذاتيا من DNMT1. يرتبط هذا المجال ، الموجود في المنطقة التنظيمية N-terminal ، بالموقع الحفاز ويمنع ارتباط الحمض النووي. تؤدي إزالة أول ~ 600 من الأحماض الأمينية إلى إنزيم مقطوع أكثر نشاطا بشكل ملحوظ من الإنزيم كامل الطول (~ زيادة 640 ضعفا في kcat / Km) 40. يسمح هذا الشكل المنشط من الإنزيم ، المشار إليه باسم DNMT1 الذي يفتقر إلى RFTS (الأحماض الأمينية 621-1616) ، بتحديد أسهل للمثبطات بسبب قوته التحفيزية المتزايدة. تقدم هذه الورقة بروتوكولا لاستخدام DNMT1 الذي يفتقر إلى RFTS في المقايسات لفحص مثبطات الجزيئات الصغيرة المحتملة. باستخدام الفحص المستمر المقترن بالنوكلياز الداخلي ، يتم تحديد السرعة الأولية في وجود وغياب عدد قليل من الجزيئات الصغيرة. يتم فحص كل مثبط محتمل بتركيزين للبحث عن تثبيط DNMT1 المعتمد على التركيز. تم حساب النسبة المئوية للنشاط الذي لوحظ في وجود الجزيئات الصغيرة في كل حالة.
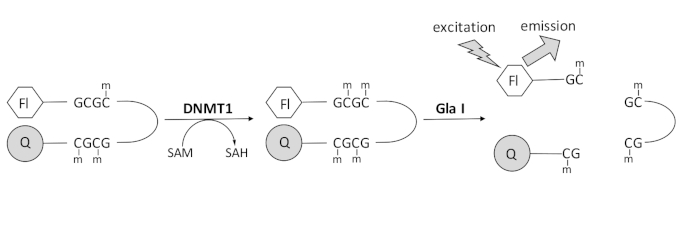
الشكل 1: مقايسة مثيلة الحمض النووي. يتم استخدام الحمض النووي لدبوس الشعر hemimethylated مع فلوروفور في نهاية 5 ‘ومطفأ على نهاية 3’ كركيزة. يحفز DNMT1 نقل مجموعة الميثيل من S-adenosylmethionine إلى موقع CpG غير الميثيل ، مما يولد S-adenosylhomocysteine والحمض النووي الميثيلي بالكامل. يحتوي منتج الحمض النووي على موقع الانقسام للنوكلياز الداخلي Gla I ، الذي يشق مواقع GCGC الميثلية بالكامل. يؤدي انشقاق الحمض النووي للمنتج إلى إطلاق الفلوروفور 5 بوصات من مخوي 3 بوصات ، مما يولد مضان. الاختصارات: Fl = فلوروفور. س = مروي. DNMT1 = الحمض النووي ميثيل ترانسفيراز 1 ؛ SAM = S-أدينوسيل ميثيونين. SAH = S-أدينوسيل هوموسيستين. الرجاء الضغط هنا لعرض نسخة أكبر من هذا الشكل.
