DNA-Methylierung ist eine wichtige epigenetische Markierung, die die Genexpression und die Chromatinstruktur reguliert. Die Methylierung erfolgt überwiegend in CpG-Dinukleotiden – Cytosin gefolgt von Guanosin; Die Methylgruppe wird zur 5-Position von Cytosin hinzugefügt. Korrekte DNA-Methylierungsmuster und damit die richtige Genexpression sind für eine angemessene zelluläre Entwicklung und Funktion erforderlich. Viele Krankheitszustände wurden mit Veränderungen des normalen Methylierungsmusters 1,2,3 in Verbindung gebracht. Zum Beispiel gibt es einen Zusammenhang zwischen Krebsentstehung und -progression und Veränderungen des DNA-Methylierungsmusters. Typischerweise weisen Krebszellen insgesamt niedrigere Methylcytosinspiegel auf, was zur Instabilität des Genoms beiträgt. Gleichzeitig konzentriert sich das im Erbgut vorhandene Methylcytosin in den Promotorregionen von Tumorsuppressorgenen, was zu einer Gen-Silencierung dieser wichtigen Proteine führt. Insbesondere epigenetische Veränderungen sind dynamisch und reversibel, im Gegensatz zu den DNA-Mutationen, die mit der Tumorgenese assoziiert sind. Dies hat die Proteine, die an der epigenetischen Genregulation beteiligt sind, zu interessantenWirkstoffzielen 2,4 gemacht.
DNA-Methyltransferasen (DNMTs) sind die Proteine, die für die Erzeugung und Aufrechterhaltung von DNA-Methylierungsmustern verantwortlich sind. Drei katalytisch aktive Isoenzyme, DNMT1, DNMT3a und DNMT3b, existieren beim Menschen. Während der Entwicklung und Differenzierung etablieren die de novo Methyltransferasen DNMT3a und DNMT3b Methylierungsmuster. Beide Enzyme können das katalytisch inaktive DNMT3L-Protein zu Komplexen binden, die eine erhöhte Aktivitätaufweisen 1,5. Nach der Zellteilung enthalten Tochterzellen hemimethylierte DNA – DNA, die Methylcytosin in nur einem Strang des Duplex enthält -, weil die neu synthetisierte DNA frei von Methylierungsmarkierungen ist. Die Hauptfunktion von DNMT1 besteht darin, diese hemimethylierte DNA zu methylieren, wodurch das vollständige Methylierungsmuster 1,5 wiederhergestellt wird.
Zusammenhänge zwischen DNMT-Aktivität und Krebs sind gut etabliert. Die Überexpression von DNMT1, entweder durch transkriptionelle oder posttranslationale Mechanismen, ist eine Folge mehrerer gemeinsamer onkogener Signalwege 6,7,8,9. Genetische Ansätze zur Senkung der DNMT1-Aktivität unter Verwendung hypomorpher Allele führen zu einer verminderten Tumorbildung bei Apc(Min)Mäusen10. Antisense-Oligonukleotide, die DNMT1 niederschlagen, hemmen Neoplasien in Zellkultur- undMaustumormodellen 11,12. Daher scheint die Hemmung der DNMT1-Aktivität ein vielversprechender Krebstherapieansatz zu sein. Die Rollen, die die DNMT3-Isoenzyme spielen, sind jedoch nicht so einfach. DNMT3a-Mutationen finden sich bei akuter myeloischer Leukämie13 und myelodysplastischem Syndrom14. Es wurde gezeigt, dass mindestens eine der identifizierten Mutationen die DNA-Methylierungsaktivität des Enzyms15 verringert. DNMT3b ist jedoch bei Brustkrebs16 und Darmkrebs17 überexprimiert. Da die verschiedenen DNMT-Isoenzyme unterschiedliche Rollen bei der Karzinogenese spielen, wird die Identifizierung isoenzymspezifischer Inhibitoren entscheidend sein. Diese Verbindungen werden nicht nur für die Entwicklung von Therapeutika nützlich sein, sondern isoenzymspezifische Inhibitoren wären auch ein unschätzbares Werkzeug, um die Rolle jedes DNMT-Isoenzyms in der Krebsätiologie zu analysieren.
Mehrere DNMT-Inhibitoren wurden in der Literatur beschrieben. Bekannte DNMT-Inhibitoren können in zwei Klassen unterteilt werden: Nukleosid und Nicht-Nukleosid. Nukleosidinhibitoren sind typischerweise Cytidinanaloga. Diese Verbindungen werden in die DNA eingebaut und fangen DNMTs kovalent ein. 5-Azacytidin und 5-Aza-2′-Desoxycytidin sind für die Behandlung des myelodysplastischen Syndroms und der akuten myeloischen Leukämie zugelassen 4,18. Die hohe Toxizität, geringe Bioverfügbarkeit und chemische Instabilität dieser Verbindungen stellen Probleme dar. Laufende Arbeiten untersuchen die Wirksamkeit der nächsten Generation von Nukleosidinhibitoren; SGI-110, abgeleitet von 5-Aza-2′-Desoxycytidin, ist ein Beispiel19,20. Nukleosid-Inhibitoren sind nicht isoenzymspezifisch und inaktivieren jedes auftretende DNMT-Isoenzym. Daher führt die Behandlung mit einem nukleosiddemethylierenden Mittel zur Erschöpfung aller DNMT-Isoenzyme 4,18. Nicht-Nukleosid-Inhibitoren müssen nicht in die DNA eingebaut werden, um ihre hemmende Wirkung auszuüben. Stattdessen binden diese Moleküle direkt an DNMTs, was die Möglichkeit einer isoenzymspezifischen Hemmung einführt. Bisher wurden mehrere Nicht-Nukleosid-Inhibitoren entdeckt, darunter SGI-1027 21, Hydralazin-22, Procainamid23, RG108 und Derivate 24 sowie die Naturstoffe (−)-Epigallocateching-3-Gallat (EGCG)25 und Milchsäure A 26,27. Die meisten der bisher entdeckten Nicht-Nukleosid-Inhibitoren sind nicht isoenzymselektiv oder zeigen schwache Präferenzen für ein DNMT-Isoenzym. Darüber hinaus muss die Wirksamkeit dieser Moleküle verbessert werden, insbesondere in Zellen 4,18. Daher besteht die Notwendigkeit, potentere, isoenzymselektive DNMT-Inhibitoren zu entdecken oder zu entwickeln.
Eine Hürde bei der Entdeckung neuer niedermolekularer Inhibitoren von DNMTs sind die aufwendigen Assays, die traditionell zur Untersuchung der DNMT-Aktivität verwendetwerden 28. Assays sind in der Regel diskontinuierlich mit mehreren Schritten. Die enzymatische Aktivität von DNMTs wird immer noch routinemäßig mit radioaktivem S-Adenosylmethionin (SAM) 29,30,31,32,33,34 untersucht. Nicht-radioaktive Assays für die DNA-Methylierung wurden ebenfalls entwickelt. Zum Beispiel wurden Assays beschrieben, die methylsensitive Restriktionsendonukleasen und Elektrophorese zur Trennung der Verdauungsprodukte verwenden35,36. Diese Arten von diskontinuierlichen, mehrstufigen Assays sind für die Wirkstoffforschung nicht ohne weiteres zugänglich. Seit Mitte der 2000er Jahre wurden mehrere DNA-Methylierungsassays mit höherem Durchsatz entwickelt28. Ein Szintillations-Proximity-Assay wurde verwendet, um nach DNMT1-Inhibitoren zu suchen37. Ein weiterer Assay unter Verwendung einer methylsensitiven Restriktionsendonuklease wurde verwendet, um nach DNMT3a-Inhibitoren25,38 zu suchen. Während beide Assays einen höheren Durchsatz als herkömmliche DNA-Methylierungsassays ermöglichten, erfordern die Assays mehrere Schritte und erlauben nicht die Beobachtung der Methylierungsaktivität in Echtzeit. In jüngerer Zeit wurde ein kontinuierlicher Kinetik beschrieben, der die Bildung von S-Adenosylhomocystein (SAH), einem Produkt der Methylierungsreaktion, an die spektroskopische Veränderung bei 340 nm koppelt, die mit der NADPH-Oxidation39 assoziiert ist. Dieser Assay verwendet drei Kopplungsenzyme, um ein spektroskopisches Signal zu erzeugen.
Wir entwickelten einen fluoreszenzbasierten Endonuklease-gekoppelten DNA-Methylierungsassay, der ein einzelnes kommerziell verfügbares Kopplungsenzym verwendet und Daten in Echtzeit generieren kann (Abbildung 1). Als Substrat wird ein Haarnadel-Oligonukleotid verwendet, das drei Methylcytosine enthält. Die Substrat-DNA enthält einen Fluorophor am 5′-Ende und einen Quencher am 3′-Ende. Die Methylierung der hemimethylierten CpG-Stelle erzeugt die Spaltstelle für die Endonuklease Gla I – vollständig methyliertes GCGC. Die Gla-I-Spaltung des Produktoligonukleotids setzt den Fluorophor aus dem Quencher frei und erzeugt Fluoreszenz in Echtzeit. Der Assay kann verwendet werden, um die Aktivität jeder Isoform von DNMT zu untersuchen; jedoch wird eine höhere Aktivität mit DNMT1 beobachtet, da dieses Isoenzym bevorzugt hemimethylierte DNA 1,5 methyliert. Eine noch robustere Aktivität wird beobachtet, wenn die autoinhibitorische RFTS-Domäne (Replication Foci Targeting Sequence) aus DNMT1 entfernt wird. Diese Domäne, die sich in der N-terminalen regulatorischen Region befindet, bindet an die katalytische Stelle und verhindert die DNA-Bindung. Die Entfernung der ersten ~600 Aminosäuren führt zu einem verkürzten Enzym, das signifikant aktiver ist als das Enzym in voller Länge (~640-facher Anstieg in kKatze / Km)40. Diese aktivierte Form des Enzyms, die als RFTS-fehlendes DNMT1 (Aminosäuren 621–1616) bezeichnet wird, ermöglicht aufgrund ihrer erhöhten katalytischen Kraft die leichtere Identifizierung von Inhibitoren. Dieses Papier stellt ein Protokoll vor, um RFTS-fehlendes DNMT1 in Assays zu verwenden, um nach potenziellen niedermolekularen Inhibitoren zu suchen. Mit dem Endonuklease-gekoppelten kontinuierlichen Assay wird die Anfangsgeschwindigkeit in Gegenwart und Abwesenheit einiger kleiner Moleküle bestimmt. Jeder potenzielle Inhibitor wird bei zwei Konzentrationen untersucht, um nach konzentrationsabhängiger DNMT1-Hemmung zu suchen. Die prozentuale Aktivität, die in Gegenwart der kleinen Moleküle beobachtet wurde, wurde jeweils berechnet.
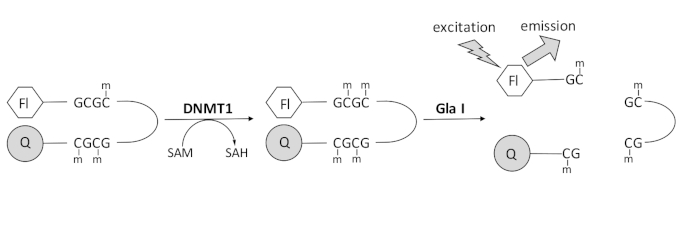
Abbildung 1: DNA-Methylierungstest. Als Substrat wird eine hemimethylierte Haarnadel-DNA mit einem Fluorophor am 5′-Ende und einem Quencher am 3′-Ende verwendet. DNMT1 katalysiert den Transfer der Methylgruppe von S-Adenosylmethionin zur nicht methylierten CpG-Stelle und erzeugt S-Adenosylhomocystein und vollständig methylierte DNA. Das DNA-Produkt enthält die Spaltstelle für die Endonuklease Gla I, die vollständig methylierte GCGC-Stellen spaltet. Die Spaltung der Produkt-DNA setzt den 5′-Fluorophor aus dem 3′-Quencher frei und erzeugt Fluoreszenz. Abkürzungen: Fl = Fluorophor; Q =Quencher; DNMT1 = DNA-Methyltransferase 1; SAM = S-Adenosylmethionin; SAH = S-Adenosylhomocystein. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
