Domates (S. lycopersicum L.), 2020 yılında 5,1 milyon hektardan 186,8 milyon ton etli meyve üretimi ile dünyadaki en önemli sebze mahsullerinden biridir1. Patlıcan, biber, patates ve tütün gibi ticari açıdan önemli birçok ürün de dahil olmak üzere yaklaşık 2.716 tür2 ile büyük Solanaceae familyasına aittir. Ekili domates, genom boyutu yaklaşık 900 Mb3 olan bir diploid türüdür (2n = 2x = 24). Uzun süredir, yabani Solanum spp’den arzu edilen özellikleri seçerek domates evcilleştirme ve ıslahına yönelik büyük çaba sarf edilmiştir. Domates Genetiği Kaynak Merkezi’nde listelenen 5.000’den fazla domates katılımı vardır ve dünya çapında 80.000’den fazla germplazma domates depolanmaktadır4. Domates bitkisi serada çok yıllıktır ve tohumlarla yayılır. Olgun bir domates tohumu üç ana bölmeden oluşur: tam yetişkin bir embriyo, artık hücresel tip endosperm ve sert bir tohum kabuğu 5,6 (Şekil 1A). Çift döllenmeden sonra, hücresel tip endosperm gelişimi, zigotların gelişiminden önce gelir. Çiçeklenmeden (DAF) ~ 5-6 gün sonra, endosperm altı ila sekiz çekirdek7’den oluştuğunda iki hücreli proembriyo ilk önce gözlenir. Solanum pimpinellifolium’da, embriyo 20 DAF’tan sonra son boyutuna yaklaşır ve tohumlar 32 DAF8’den sonra çimlenme için uygundur. Embriyo geliştikçe, endosperm yavaş yavaş emilir ve tohumda sadece az miktarda endosperm kalır. Kalıntı endosperm, radikül ucunu çevreleyen mikropylar endosperm ve tohumun geri kalanında lateral endosperm 9,10’dan oluşur. Dış tohum kabuğu, bütünlüğün kalınlaşmış ve odunlaşmış dış epidermisinden gelişir ve bütünleşme kalıntılarının ölü katmanlarıyla, embriyoyu ve endospermi korumak için sert bir kabuk oluştururlar5.
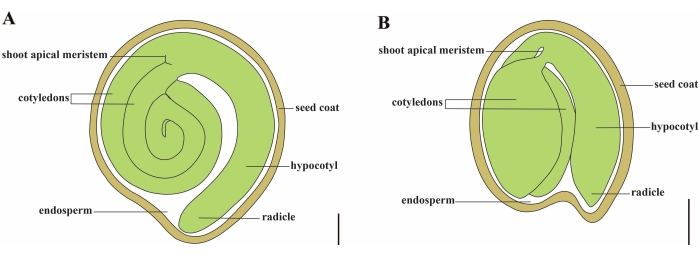
Resim 1: Solanum lycopersicum ve Arabidopsis thaliana’da olgun bir tohumun şematik gösterimi . (A) Olgun bir domates tohumunun uzunlamasına anatomisi. (B) Olgun bir Arabidopsis tohumunun uzunlamasına anatomisi. Olgun bir domates tohumu, bir Arabidopsis tohumundan yaklaşık 70 kat daha büyüktür. Ölçek çubukları = (A) 400 μm, (B) 100 μm. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
Yüksek kaliteli domates tohumlarının üretimi, embriyo, endosperm ve maternal tohum bileşenleri arasındaki koordinasyona bağlıdır11. Tohum gelişimindeki anahtar genlerin ve ağların diseksiyonu, mutant tohumların derin ve tam hatlı bir fenotipik kaydını gerektirir. Yarı ince kesit ve parafin kesit gibi geleneksel gömme-kesit teknikleri, embriyonun yerel ve daha ince yapılarını gözlemlemek için domates tohumlarına yaygın olarak uygulanır12,13,14,15. Bununla birlikte, tohum gelişimini ince kesitlerden analiz etmek genellikle zahmetlidir ve z ekseni uzamsal çözünürlüğünden yoksundur. Buna karşılık, doku temizleme, ortaya çıkması en muhtemel embriyo defektlerinin gelişim aşamasını belirlemek için hızlı ve etkili bir yöntemdir16. Temizleme yöntemi, kırılma indisini bir veya daha fazla biyokimyasal ajanla homojenleştirerek iç dokunun opaklığını azaltır16. Tüm doku temizleme, bir bitki doku yapısının bütünlüğünü bozmadan gözlemlenmesine izin verir ve temizleme teknolojisi ile üç boyutlu görüntülemenin kombinasyonu, bir bitki organının morfolojisi ve gelişimsel durumu hakkında bilgi edinmek için ideal bir çözüm haline gelmiştir17,18. Yıllar geçtikçe, Arabidopsis thaliana, Hordeum vulgare ve Beta vulgaris19,20,21,22,23 dahil olmak üzere çeşitli bitki türlerinde tohum temizleme teknikleri kullanılmıştır. Bunlar arasında, bütün monte edilmiş ovül temizleme teknolojisi, küçük boyutu, tohum kabuğu hücresinin 4-5 katmanı ve nükleer tip endosperm24,25 nedeniyle Arabidopsis’in tohum gelişimini incelemek için etkili bir yaklaşım olmuştur. Hoyer çözeltisi26’nın ortaya çıkması gibi farklı temizleme karışımlarının sürekli güncellenmesiyle, arpa ovülünün iç yapıları, endospermi tohumların büyük kısmını oluşturmasına rağmen, yüksek derecede netlikle görüntülenmiştir. Şeker pancarının embriyogenezi, vakum işlemi ile birlikte temizlenerek ve hidroklorik asit19 ile yumuşatılarak gözlemlenebilir. Bununla birlikte, yukarıda belirtilen türlerin aksine, domates tohumlarındaki protokollerin temizlenmesiyle embriyolojik gözlemler bildirilmemiştir. Bu, domateslerin embriyonik ve tohum gelişiminin ayrıntılı bir şekilde araştırılmasını önler.
Kloral hidrat, daldırılmış dokuların ve hücrelerin farklı optik düzlemlerde görüntülenmesini sağlayan ve hücreleri veya doku bileşenlerini büyük ölçüde koruyan bir temizleme çözeltisi olarak yaygın olarak kullanılır27,28,29. Kloral hidrat bazlı temizleme protokolü, Arabidopsis21,28’in embriyosunu ve endospermini gözlemlemek için tohumların tamamen temizlenmesi için başarıyla kullanılmıştır. Bununla birlikte, bu temizleme çözeltisi, Arabidopsis tohumlarından daha geçirimsiz olan domates tohumlarının temizlenmesinde etkili değildir. Fiziksel engeller şunları içerir: (1) domates bütünlüğünün 3 ila 15 DAF 30,31’de yaklaşık 20 hücre katmanı vardır, (2) domates endospermi nükleer tip 32 değil, hücresel tiptedir ve (3) domates tohumları 33,34 boyutunda yaklaşık70 kat daha büyüktür ve (4) temizleme reaktiflerinin penetrasyonunu engelleyen ve embriyo hücrelerinin görselleştirilmesini etkileyen büyük miktarlarda tohum kabuğu müsilajı üretir.
Bu nedenle, bu rapor, domates tohumlarının farklı aşamalarda tamamen monte edilmesi için optimize edilmiş bir kloral hidrat bazlı temizleme yöntemi sunar ve bu da embriyo gelişim sürecinin derin görüntülenmesini sağlar (Şekil 2).
