Tomat (S. lycopersicum L.) er en av de viktigste vegetabilske avlingene rundt om i verden, med en produksjon på 186,8 millioner tonn kjøttfulle frukter fra 5,1 millioner hektar i 20201. Den tilhører den store Solanaceae-familien med ca 2,716 arter2, inkludert mange kommersielt viktige avlinger som aubergine, paprika, potet og tobakk. Den kultiverte tomaten er en diploid art (2n = 2x = 24) med en genomstørrelse på ca. 900 Mb3. I lang tid har det blitt gjort store anstrengelser mot tomatdomestisering og avl ved å velge ønskelige egenskaper fra vill Solanum spp. Det er over 5000 tomattilganger oppført i Tomato Genetics Resource Center, og mer enn 80.000 bakterieplasma av tomater lagres over hele verden4. Tomatplanten er flerårig i drivhuset og forplantes av frø. Et modent tomatfrø består av tre hovedrom: et fullvoksent embryo, gjenværende endosperm av cellulær type og et hardt frøbelegg 5,6 (figur 1A). Etter dobbel befruktning går utviklingen av cellulær endosperm foran utviklingen av zygoter. Ved ~ 5-6 dager etter blomstring (DAF) observeres tocellet proembryo først når endospermen består av seks til åtte kjerner7. I Solanum pimpinellifolium nærmer embryoet seg sin endelige størrelse etter 20 DAF, og frø er levedyktige for spiring etter 32 DAF8. Etter hvert som embryoet utvikler seg, absorberes endospermen gradvis, og bare en liten mengde endosperm forblir i frøet. Den gjenværende endospermen består av mikropylær endosperm som omgir radikelspissen, og lateral endosperm i resten av frøet 9,10. Den ytre frøbelegget er utviklet fra fortykket og lignifisert ytre epidermis av integumentet, og med de døde lagene av integumentrester danner de et hardt skall for å beskytte embryoet og endospermen5.
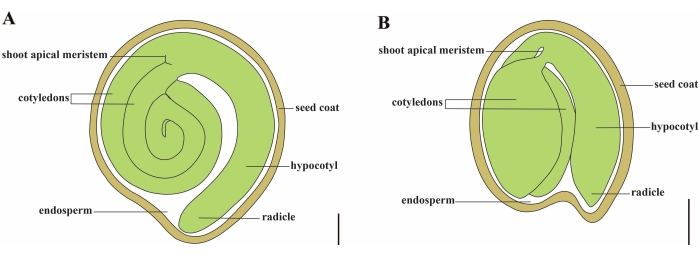
Figur 1: Skjematisk fremstilling av et modent frø i Solanum lycopersicum og Arabidopsis thaliana. (A) Langsgående anatomi av et modent tomatfrø. (B) Langsgående anatomi av et modent Arabidopsis-frø. Et modent tomatfrø er omtrent 70 ganger større i størrelse enn et Arabidopsis-frø. Skala barer = (A) 400 μm, (B) 100 μm. Vennligst klikk her for å se en større versjon av dette tallet.
Produksjon av tomatfrø av høy kvalitet avhenger av koordineringen mellom embryoet, endospermen og morsfrøkomponentene11. Dissekering av viktige gener og nettverk i frøutvikling krever en dyp og fullsporet fenotypisk registrering av mutante frø. Konvensjonelle innebyggingsteknikker, som den halvtynne delen og parafinseksjonen, brukes mye på tomatfrø for å observere de lokale og finere strukturene til embryoet12,13,14,15. Imidlertid er det vanligvis arbeidskrevende å analysere frøutviklingen fra tynne seksjoner og mangler romlig oppløsning på z-aksen. Til sammenligning er vevsrydding en rask og effektiv metode for å finne utviklingsstadiet av embryodefekter som mest sannsynlig vil oppstå16. Clearingmetoden reduserer ugjennomsiktigheten av indre vev ved å homogenisere brytningsindeksen med ett eller flere biokjemiske midler16. Hele vevsrydding tillater observasjon av en plantevevstruktur uten å ødelegge integriteten, og kombinasjonen av clearingteknologi og tredimensjonal avbildning har blitt en ideell løsning for å få informasjon om morfologien og utviklingstilstanden til et planteorgan17,18. Gjennom årene har frøryddingsteknikker blitt brukt i forskjellige plantearter, inkludert Arabidopsis thaliana, Hordeum vulgare og Beta vulgaris 19,20,21,22,23. Blant disse har hele eggløsningsryddingsteknologien vært en effektiv tilnærming til å studere frøutvikling av Arabidopsis, på grunn av sin lille størrelse, 4-5 lag av frøbeleggcellen og endospermen av kjernetypen24,25. Med kontinuerlig oppdatering av forskjellige clearingblandinger, for eksempel fremveksten av Hoyers løsning26, ble indre strukturer av byggeggløsningen avbildet med høy grad av klarhet, selv om endospermen utgjør hoveddelen av frøene. Embryogenese av sukkerroer kan observeres ved rydding kombinert med vakuumbehandling og mykning med saltsyre19. Ikke desto mindre, i motsetning til artene nevnt ovenfor, har embryologiske observasjoner ved å rydde protokoller i tomatfrø ikke blitt rapportert. Dette forhindrer detaljert undersøkelse av embryonal og frøutvikling av tomater.
Kloralhydrat brukes ofte som en clearingløsning som gjør at nedsenket vev og celler kan vises på forskjellige optiske plan, og bevarer cellene eller vevskomponentene vesentlig27,28,29. Kloralhydratbasert clearingprotokoll har blitt brukt til helmontering av frø for å observere embryo og endosperm av Arabidopsis21,28. Denne clearingløsningen er imidlertid ikke effektiv når det gjelder å rydde tomatfrø, som er mer ugjennomtrengelige enn Arabidopsisfrø. De fysiske barrierene inkluderer: (1) tomatintegumentet har nesten 20 cellelag ved 3 til 15 DAF 30,31, (2) tomatendospermen er cellulær type, ikke nukleær-type32, og (3) tomatfrø er omtrent 70 ganger større i størrelse33,34 og (4) produserer store mengder frøbelegg, som blokkerer penetrasjonen av clearingreagenser og påvirker visualiseringen av embryoceller.
Derfor presenterer denne rapporten en optimalisert kloralhydratbasert clearingmetode for helmontering av tomatfrø på forskjellige stadier, noe som muliggjør dyp avbildning av embryoutviklingsprosessen (figur 2).
