G-eiwit-gekoppelde receptoren (GPCR’s), die aanwezig zijn van gist tot mens, vertegenwoordigen de grootste superfamilie van receptoren in veel organismen1. Ze spelen een cruciale rol bij het reguleren van bijna alle biologische processen bij dieren. Er zijn 50-200 GPCR’s in het genoom van geleedpotigen, wat betekent dat ze de grootste membraanreceptor superfamilie2 vertegenwoordigen. Ze worden ingedeeld in zes hoofdklassen, A-F, op basis van hun volgordeovereenkomst en functies3. GPCR’s transduceren verschillende extracellulaire signalen, zoals die van hormonen, neuropeptiden, biogene amines, glutamaat, proton, lipoglycoproteïnen en fotonen4. GPCR’s koppelen aan heterotrimeer G-eiwitten (Gα, Gβ en Gγ) om stroomafwaartse signalen te verzenden. GPCR’s gekoppeld aan Gαs– of Gαi/o-eiwitten verhogen of verlagen respectievelijk de intracellulaire 3′, 5′-cyclische adenosinemonofosfaat (cAMP) niveaus door adenylylcyclase te activeren of te remmen. GPCR’s gekoppeld aan Gαq/11 induceren calciumafgifte uit de endoplasmatische reticulumcalciumvoorraden door de fosfolipase C (PLC)-inositol-1,4,5-trifosfaat (IP3) route te activeren. GPCR’s gekoppeld aan Gα12/13 activeren RhoGTPase nucleotide-uitwisselingsfactoren 5,6. GPCR’s zijn het doelwit van meer dan 50% van de menselijke geneesmiddelen en een acaricide, amitraz4. Omdat GPCR’s zulke uiteenlopende signalen transduceren, zijn ze veelbelovende doelen voor het ontwikkelen van nieuwe pesticiden die ongewervelde fysiologische functies verstoren.
Het doel van HTS is om hitmoleculen te identificeren die receptorfuncties kunnen moduleren. HTS omvat assay-ontwikkeling, miniaturisatie en automatisering7. Geleedpotige neuropeptide GPCRRs zijn betrokken bij de meeste fysiologische functies, zoals ontwikkeling, rui en ecdysis, uitscheiding, energiemobilisatie en reproductie4. De meeste neuropeptide GPCRRs van geleedpotigen en metazoën signaleren via de calciumsignaleringscascade 2,6,8,9,10, zoals in de myoinhibitory peptide en SIFamide receptoren van de zwartpootteek Ixodes scapularis; hun liganden zijn antagonistisch in hindgutmotiliteitstests, waarbij SIF contractie uitlokt en MIP hetremt 11,12. Een NPY-achtige receptor van de gelekoortsmug, Aedes aegypti, reguleert de vrouwelijke gastheer op zoek naar13. In vergelijking met andere alternatieve calciummobilisatietests zoals de aequorinecalciumbioluminescentietest14, is de calciumfluorescentietest gemakkelijk uit te voeren, vereist geen transfectie van andere recombinante calciumdetectie-eiwitten en is het kosteneffectief. De calciumfluorescentietest produceert een langdurig signaal in vergelijking met het snelle kinetische signaal dat wordt verkregen in de aequorinecalciumbioluminescentietest14,15.
In het voorbeeld hier werd de kininereceptor van de runderkoortsteek, Rhipicephalus microplus, recombinant tot expressie gebracht in de CHO-K1-cellijn en gebruikt voor de calciumfluorescentietest. Er is slechts één kininereceptorgen gevonden in R. microplus; de receptor signaleert via een Gq-eiwitafhankelijke signaalroute en activeert de efflux van Ca2+ uit calciumvoorraden naar de intracellulaire ruimte16. Dit proces kan worden gedetecteerd en gekwantificeerd door een fluorofoor, die een fluorescentiesignaal opwekt bij het binden van calciumionen (figuur 1).
De kininereceptor is een ongewervelde GPCR, die behoort tot de klasse A Rhodopsine-achtige receptoren. Kinine is een oud signaal neuropeptide dat aanwezig is in Mollusca, Crustacea, Insecta en Acari 4,17,18. Coleopteranen (kevers) missen het kininesignaleringssysteem; in de mug Aedes aegypti is er slechts één kininereceptor die drie aedeskinines bindt, terwijl Drosophila melanogaster één kininereceptor heeft met drosokinine als een uniek ligand 19,20,21. Er zijn geen homologe kininen of kininereceptoren in gewervelde dieren. Hoewel de exacte functie van kinine onbekend is bij teken, vertonen de kininereceptor RNAi-gedempte vrouwtjes van R. microplus een significant verminderde reproductieve fitheid22. Kininen zijn pleotrope peptiden in insecten. Bij Drosophila melanogaster zijn ze betrokken bij zowel het centrale als het perifere zenuwstelsel23, pre-ecdysis24, voeding25, metabolisme26 en slaapactiviteitspatronen26,27, evenals larvale voortbeweging28. Kinins reguleren hindgut contractie, diurese en voeding in de mug A. aegypti 29,30,31. De kininepeptiden hebben een geconserveerd C-terminal pentapeptide Phe-X1-X2-Trp-Gly-NH2, wat de minimaal vereiste sequentie is voor biologische activiteit32. De geleedpotige specificiteit, de kleine omvang van het endogene ligand, waardoor ze vatbaar zijn voor interferentie met kleine moleculen, en de pleiotrope functies bij insecten maken de kininereceptor een veelbelovend doelwit voor ongediertebestrijding4.
De “dual-addition” assay (Figuur 2) maakt de identificatie van agonisten of antagonisten in dezelfde HTS assay15 mogelijk. Het is aangepast van een “dual-addition” assay die vaak wordt gebruikt in de farmaceutische industrie voor het ontdekken van geneesmiddelen33. Kortom, de eerste toevoeging van geneesmiddelen aan de celplaat maakt de identificatie van potentiële agonisten in de chemische bibliotheek mogelijk wanneer een hoger fluorescentiesignaal wordt gedetecteerd in vergelijking met de toepassing van de oplosmiddelcontrole. Na 5 minuten incubatie met deze kleine moleculen wordt een bekende agonist (kininepeptide) op alle putten aangebracht. Die putten die willekeurig een antagonist van de medicijnplaat ontvingen, vertonen een lager fluorescentiesignaal bij agonisttoevoeging in vergelijking met de controleputten die het oplosmiddel in de eerste toevoeging ontvingen. Deze test maakt het vervolgens mogelijk om potentiële agonisten en antagonisten met dezelfde cellen te identificeren. In een standaard HTS-project zouden deze hitmoleculen verder worden gevalideerd door middel van dosis-responstests en door aanvullende biologische activiteitstests, die hier niet worden getoond.
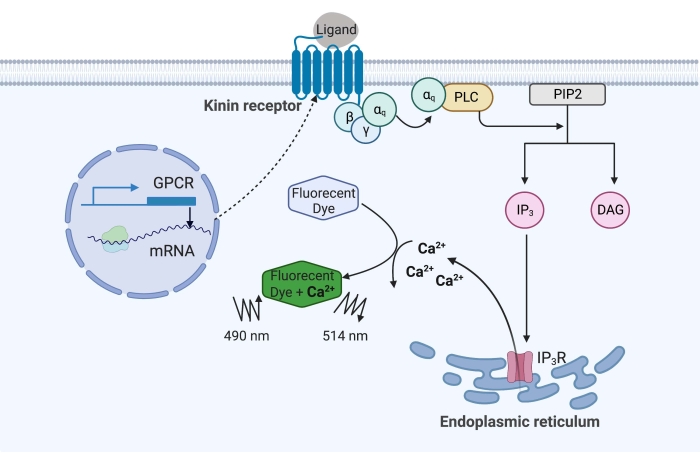
Figuur 1: Illustratie van het calciumfluorescentietestmechanisme. Het Gq-eiwit activeert de intracellulaire calciumsignaleringsroute. De kininereceptor (G-eiwit-gekoppelde receptor) werd recombinant tot expressie gebracht in CHO-K1-cellen. Wanneer de agonistligand zich aan de receptor bindt, activeert het Gq-eiwit geassocieerd met de kininereceptor PLC, dat de omzetting van een PIP 2-molecuul in IP3 en DAG katalyseert. IP3 bindt vervolgens aan de IP3R op het oppervlak van het endoplasmatisch reticulum, wat leidt tot de afgifte van Ca2+ in het cytoplasma, waar Ca2+ ionen binden aan de fluoroforen en een fluorescentiesignaal opwekken. Het fluorescentiesignaal kan worden verkregen door excitatie bij 490 nm en gedetecteerd bij 514 nm. Afkortingen: GPCR = G eiwit-gekoppelde receptor; PLC = fosfolipase C; PIP2 = fosfatidylinositol 4,5-bisfosfaat; IP3 = inositol trisfosfaat; DAG = diacylglycerol; IP3R = IP3 receptor. Gemaakt met BioRender.com. Klik hier om een grotere versie van deze figuur te bekijken.
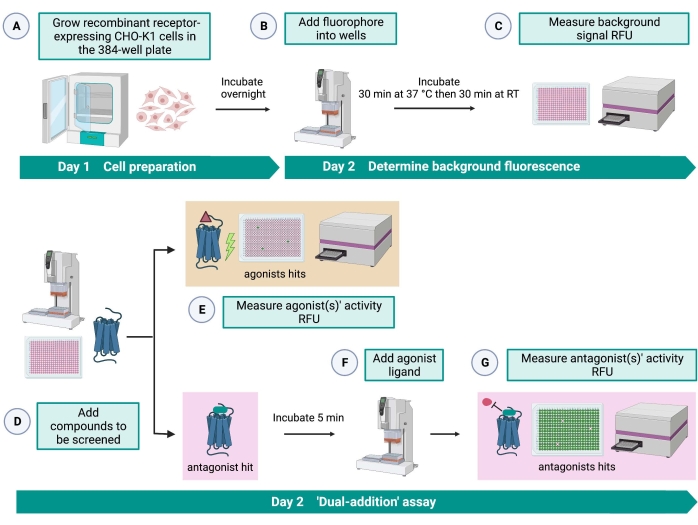
Figuur 2: De workflow voor de high-throughput screening van kleine moleculen op een G-eiwit gekoppelde receptor tot expressie gebracht in CHO-K1 cellen. (A) Recombinante CHO-K1 cellen die stabiel de kininereceptor tot expressie brengen, werden toegevoegd aan de 384-well plaat (10.000 cellen / put) met behulp van een vloeistofbehandelingssysteem (25 μL / put) en geïncubeerd in een bevochtigde CO2-incubator gedurende 12-16 uur. (B ) De testbuffer met de fluorescerende kleurstof (25 μL/put) werd met behulp van een vloeistofbehandelingssysteem aan de celplaat toegevoegd. De plaat werd gedurende 30 minuten geïncubeerd bij 37 °C gedurende 30 minuten en bij RT gedurende nog eens 30 minuten in evenwicht gebracht. (C) Het achtergrondfluorescentiesignaal van de cellen in elke put werd gemeten met een platenlezer. (D) Geneesmiddeloplossingen van een 384-well bibliotheekplaat en blanco oplosmiddel (allemaal bij 0,5 μL / put) werden toegevoegd aan de cellulaire testplaat met behulp van een vloeistofbehandelingssysteem. (E) Cellulaire calciumfluorescentieresponsen werden gemeten met de platenlezer onmiddellijk na de toevoeging van de geneesmiddeloplossingen; verbinding(en) die hoger dan gemiddelde fluorescentiesignalen uitlokken, werden uitgezocht als agonistische hit(s). Antagonist-hits die de GPCR blokkeren (pictogram hieronder) werden onthuld na de toevoeging van de peptide-agonist tijdens stap G. (F) In dezelfde testplaat werd na 5 minuten incubatie van de cellen met screeningverbindingen een endogene agonist peptide Rhimi-K-1 (QFSPWGamide) van de tekenkininereceptor aan elke put toegevoegd (1 μM). (G) Cellulaire fluorescentiereacties na de toevoeging van het agonistpeptide werden onmiddellijk door de plaatlezer gemeten. Verbinding(en) die het fluorescentiesignaal remmen werden geselecteerd als antagonist hit(s). Afkortingen: GPCR = G eiwit-gekoppelde receptor; RT = kamertemperatuur; RFU = relatieve fluorescentie-eenheden. Gemaakt met BioRender.com. Klik hier om een grotere versie van deze figuur te bekijken.
