The immunoblotting activity of the non-adsorbed polyclonal Mycoplasma pneumoniae antiserum to heterologous bacteria
Cross-reactivity does indeed exist as showcased in the immunoblotting results (Figure 2) and compared to the positive control (Lane 1), some of the M. pneumoniae antigens are shared with the screened bacteria. The intensity of these cross-reactions was variable. For instance, M. gallisepticum and M. imitans antigens displayed the strongest reactivity with M. pneumoniae antiserum (Lanes 9 and 10), besides being avian Mycoplasmas. Opposingly, neither of the two human clinical isolates of Ureaplasma urealyticum showed any reactivity (Lanes 7 and 8). However, antigens of the remaining human genital Mycoplasma species M. hominis, M. fermentans, and M. genitalium yielded considerable cross-reactions with M. pneumoniae antiserum (Lanes 11, 12, and 13, respectively). Escherichia coli (Lane 2), Pseudomonas aeruginosa (Lane 4), and Klebsiella pneumoniae (Lane 6) also reacted with polyclonal M. pneumoniae antibodies, and their antigenic profile pattern was very similar. This profile was slightly different from those of the two cocci bacteria species Streptococcus pneumoniae and Staphylococcusaureus; the reactivity was less with S. pneumoniae (Lane 3) and even lesser with S.aureus (Lane 5).
Specificity of Mycoplasma pneumoniae antibodies guaranteed by adsorption procedure
After the adsorption of M. pneumoniae polyclonal antiserum against the heterologous bacterial antigens, the specificity was tested by immunoblotting (Figure 3). Apart from some proteins of M. pneumoniae antigen revealed in Lane 1 (positive control), the other antigens were almost completely non-detected (Lanes 2-13). The revealed proteins in Lane 1 are actually the specific proteins detected by the specific M. pneumoniae antibodies remaining after adsorption. Based on this result, the efficiency of the adsorption procedure was proved, since the encountered nonspecific reactions of M. pneumoniae polyclonal antiserum with the heterologous antigens, described above, were eliminated. M. pneumoniae antiserum rendered specific was then used in all the subsequent serological and immunoblotting tests.
Capture-antigen ELISA: Valid assay for routine laboratory serodiagnostic activity
Since all sera were tested in duplicate wells for the different dilutions, OD mean values were calculated and a bar graph correlating these absorbance values to their corresponding serum dilutions was plotted (Figure 4). Based on IP calculation, the set of human sera tested and presented in this paper proved positive for the M. pneumoniae IgG. Several other serum sets were also tested using this homemade ELISA, and at each time the assay proved efficient in specifically detecting the M. pneumoniae IgG and thus distinguishing the M. pneumoniae-infected patients from those uninfected (data not shown).
ELISA results were always confirmed by several immunoblot analyses simultaneously showing the positivity in M. pneumoniae and the negativity in all the other bacteria of any tested serum found to be positive by the in-house capture-antigen M. pneumoniae ELISA (data not shown).
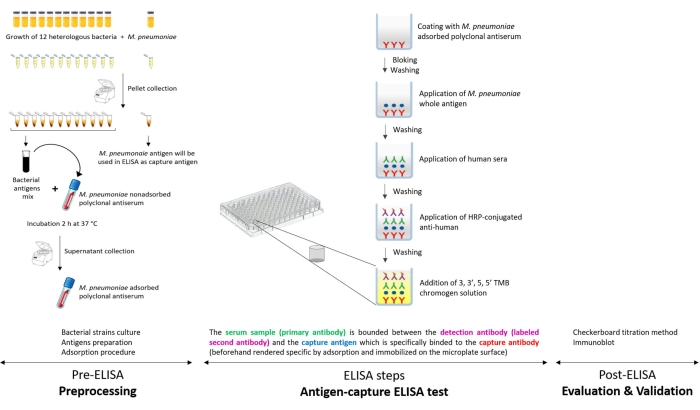
Figure 1: Schematic presentation of the antigen-capture ELISA developed in the laboratory for specific detection of Mycoplasma pneumoniae IgG. Please click here to view a larger version of this figure.
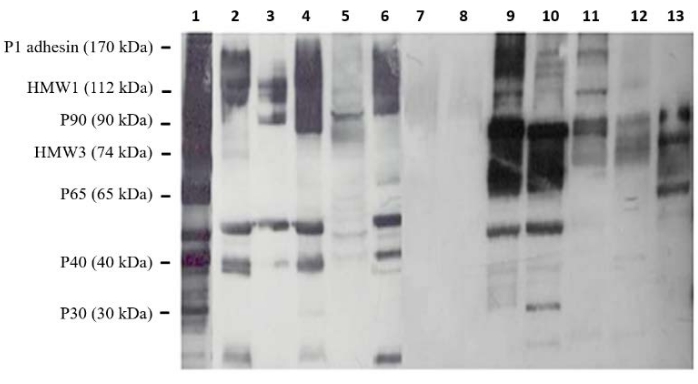
Figure 2: Immunoblot analysis of cross-reactions between Mycoplasma pneumoniae and a set of heterologous bacteria. The reactivity of M. pneumoniae non-adsorbed polyclonal antiserum was tested against antigens of Escherichia coli (Lane 2), Streptococcus pneumoniae (Lane 3), Pseudomonas aeruginosa (Lane 4), Staphylococcus aureus (Lane 5), Klebsiella pneumoniae (Lane 6), two Ureaplasma urealyticum isolates (Lanes 7 and 8), M. imitans (Lane 9), M. gallisepticum (Lane 10), M. hominis (Lane 11), M. fermentans (Lane 12), and M. genitalium (Lane 13). Lane 1: M. pneumoniae (positive control). Molecular weights of some major M. pneumoniae proteins were given on the left. Please click here to view a larger version of this figure.

Figure 3: Immunoblot confirmation of the efficiency of the adsorption procedure to eliminate cross-reactions. Reactivity of all the selected heterologous bacteria; Escherichia coli, Streptococcus pneumoniae, Pseudomonas aeruginosa, Staphylococcus aureus, Klebsiella pneumoniae, two Ureaplasma urealyticum isolates, M. imitans, M. gallisepticum, M. hominis, M. fermentans, and M. genitalium (Lanes 2-13, respectively) was tested against the adsorbed M. pneumoniae polyclonal antiserum. M. pneumoniae antigen was loaded in Lane 1 as positive control (only specific proteins were revealed). Lane MW: pre-stained protein ladder. Please click here to view a larger version of this figure.
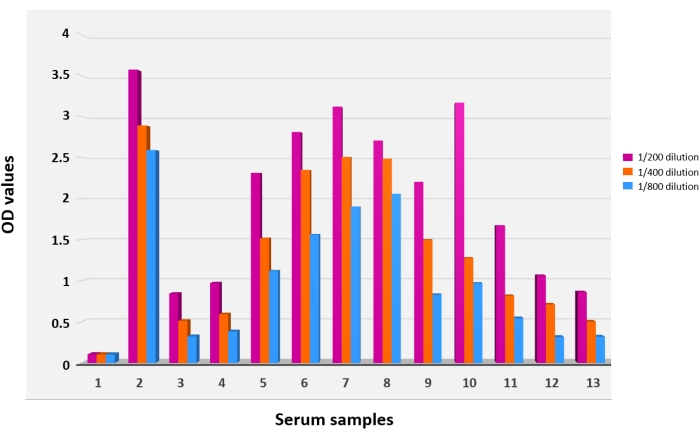
Figure 4: Results of the antigen-capture ELISA. The ELISA was performed using the adsorbed Mycoplasma pneumoniae polyclonal antiserum as a coating agent. Bar graphs 1 and 2 depict negative and positive controls, respectively. Bar graphs 3-13 depict the sera of a group of tested patients. Each serum was tested at three different dilutions: 1/200 (purple bars), 1/400 (orange bars), and 1/800 (blue bars). OD values served to calculate the index of positivity. Each reaction was performed in duplicate. Please click here to view a larger version of this figure.
| Microorganism | Strain* | Medium | Reference | |
| Mycoplasma and ureaplasma species | Mycoplasma pneumoniae | ATCC 160 20030 | SP4 | [35] |
| Mycoplasma genitalium | CIP 103767T | SP4 | ||
| Mycoplasma fermentans | ATCC 160 20026 | SP4 | ||
| Mycoplasma hominis | CIP 103715T | SP4 supplemented with arginine | ||
| Ureaplasma urealyticum | 2 clinical isolates** | SP4 supplemented with urea | ||
| Mycoplasma gallisepticum | CIP S6 15302 | Frey | [36] | |
| Mycoplasma imitans | ATCC 160 20037 | Frey | ||
| Other bacteria | Escherichia coli | ATCC 25922 | LB broth | [37] |
| Klebsiella pneumoniae | Clinical isolate*** | LB broth | ||
| Pseudomonas aeruginosa | ATCC 27853 | LB broth | ||
| Staphylococcus aureus | ATCC 25923 | LB broth | ||
| Streptococcus pneumoniae | Clinical isolate*** | Blood broth | ||
| * ATCC: American Type Culture Collection, CIP: Collection Institut Pasteur | ||||
| ** Isolates were collected from clinical samples voluntarily shipped to the Laboratory of Mycoplasmas (Pasteur Institute of Tunis) for routine diagnostic | ||||
| *** Isolates were collected from clinical samples voluntarily shipped to the Laboratory of Bacteriology (Pasteur Institute of Tunis) for routine diagnostic | ||||
Table 1: List of bacteria strains used in this study.
