Oxygenic photosynthesis is one of the most important chemical reactions on our planet. The conversion of light to chemical energy occurs in the reaction centers of two photosystems, photosystem I (PSI) and photosystem II (PSII)1 (Figure 1A). PSI is a large, highly conserved multisubunit pigment-protein complex that evolved over 3.5 billion years ago2,3. This complex, which contains approximately 100 chlorophyll molecules and about 20 carotenoids, facilitates the transfer of electrons across the thylakoid membrane from plastocyanin to ferredoxin acting as the terminal electron acceptor of the photosynthetic electron transport chain1,4,5(Figure 1B, C). In plants, this light-driven charge separation is the result of light energy transferred from both PSI core antenna pigments and the peripheral antenna pigments of Light Harvesting complex I (LHCI) to the PSI reaction center (Figure 1D). LHCI is a PSI-specific antenna complex within the thylakoid membrane composed of four chlorophyll a/b binding LHCA antenna proteins6,7.
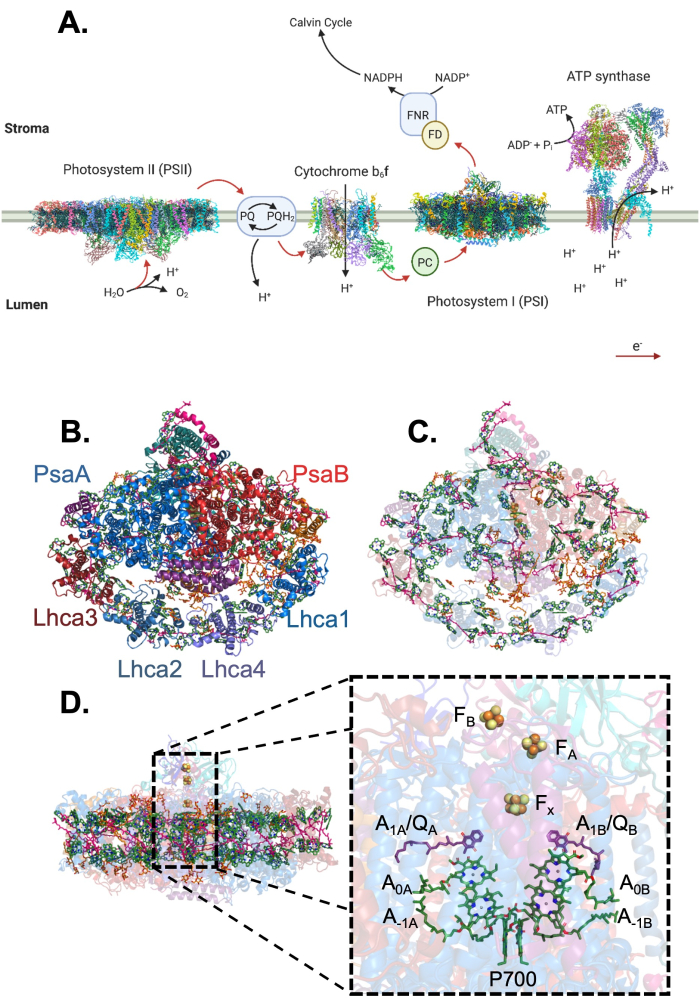
Figure 1: The photosynthetic electron transport chain and the overall structure of the PSI-LHCI complex. (A) The photosynthetic electron transport chain contains four main membrane-bound photosynthetic complexes and three soluble electron carriers. Electron flow (red arrows) through the transport chain and proton pumping (black arrows) into the lumen are used to create reducing power (NADPH) and produce ATP for carbon fixation37,38,39,40. Created with Biorender.com. (B) The structure of the plant PSI-LHCI from the lumenal side. PsaA and PsaB are the largest subunits of PSI and comprise the core of the complex. LHCI is the light-harvesting antenna complex associated with PSI and is composed of four antennae, LHCA1-4. (C) The PSI-LHCI complex coordinates over 150 ligands. Shown here are chlorophylls (green), carotenoids (pink), quinones (purple), lipids (orange), and the FeS clusters of the reaction center in yellow/orange. (D) The reaction center of PSI is split into two branches (A and B), starting from P700, the reaction center special chlorophyll pair, going into two accessory chlorophylls (A-1A/B) followed by another pair of chlorophylls (A0A/B). These chlorophylls are followed by a phylloquinone (A1A/B or QA/B in some publications) in each branch before joining together at the iron-sulfur cluster Fx followed by two more clusters, FA and FB, coordinated by the PsaC subunit. Please click here to view a larger version of this figure.
The first isolation of PSI from plants in 1966 shed light on the differences in light-harvesting pigment content between PSI and PSII, showing that PSI was highly enriched in β-carotene relative to PSII and that cytochromes f and b6 (part of the cytochrome b6f complexes) are not tightly bound to PSI but loosely associated within the thylakoid membrane8. Nine years later, with partial denaturation of isolated PSI via SDS treatment it was shown that dissociation of small PSI subunits quenched NADP+ photoreduction by PSI, while the P700 signal and most of the chlorophylls remained within the remaining large molecular weight PSI particle, identifying the necessity of some of PSI's small subunits for full biological function and the location of the PSI reaction center9. Research into the association between the PSI core and LHCI was first published in the early 1980s, when isolations of different-sized PSI species containing differing ratios of chlorophyll A to P700 were observed, suggesting the association of PSI with a chlorophyll-containing peripheral antenna system10,11,12,13. However, it wasn't until 2003 that the first crystal structure of the plant PSI was published14. The crystal structure of the plant PSI-LHCI highlighted the remarkable conservation between the PSI core of plants and cyanobacteria and provided the first picture of chlorophyll arrangement within the plant PSI core and LHCI antenna, furthering the understanding of the energy transfer pathways within the plant PSI-LHCI complex14. Over the past decade more plant PSI-LHCI structures were determined adding atomic levels details to the structural description of the super-complex15,16,17,18,19.
PSI not only has a quantum efficiency close to one, but boasts the most negative reduction potential in nature20,21. A complete understanding of PSI-LHCI and its properties is essential for understanding light driven energy transfer and applying bio-inspired solutions to future light harvesting technology. To further this understanding of how PSI-LHCI and its many subunits can achieve such efficient energy conversion, complexes isolated for study must be active and whole. This protocol allows for the gentle purification of the complex in this active state22,23.
In this method plant tissues are mechanically disrupted and chloroplasts containing the photosynthetic electron transport chain are isolated by centrifugation. The thylakoid membranes are separated after hypotonic chloroplast lysis and are then solubilized using the detergent n-dodecyl-beta-maltoside (β-DDM). The solubilized chlorophyll containing membrane complexes are separated using anion exchange chromatography and PSI-LHCI is further separated using sucrose gradient centrifugation. After removal from the gradient and after characterization by both spectroscopy and using sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), the complex can be prepared for further experiments. This procedure is used to purify the PSI-LHCI complex from plants without the use of any affinity tags. With minor modifications it can be adapted for preparations of the complex from other organisms, stabilize alternative PSI complexes or other complexes of the photosynthetic electron transport chain. Similar protocols were used to obtain PSI complex suitable for high resolution structural analysis23,24,25,26,27,28,29,30.
