With this protocol, we achieved successful isolation and dissociation of entire intestines from 5 dpf larvae. Using papain as the dissociation enzyme, we significantly enhanced cell viability, enabling the capture of 46,139 events involving single, viable cells (6.4% of all cells) out of 244 isolated guts (Figure 2A). Wildtype whole larvae were used as a control to ensure that the sorting process was optimized, enabling effective cell identification and sorting. Whole larvae could be used as we only sort for live, single cells (Supplementary Figure S1B-E). All sorted intestinal cells were subsequently submitted to the scRNA seq platform. In total, 9,858 cells were sequenced with mean reads per cell of 21,106. For scRNA-seq analysis, we used Seurat V317. In total, 48 clusters were identified representing 12 different cell types, including epithelial, stromal, blood, muscle, and immune cells, as well as enteric neurons and glia18.
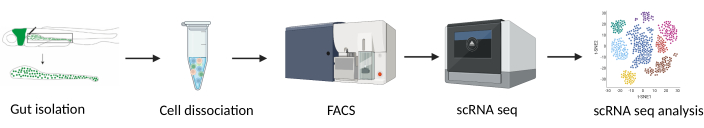
Figure 1: Experimental design for single-cell RNA sequencing of isolated zebrafish guts. Abbreviations: FACS = fluorescence-activated cell sorting; scRNA-seq = single-cell RNA-sequencing. Please click here to view a larger version of this figure.
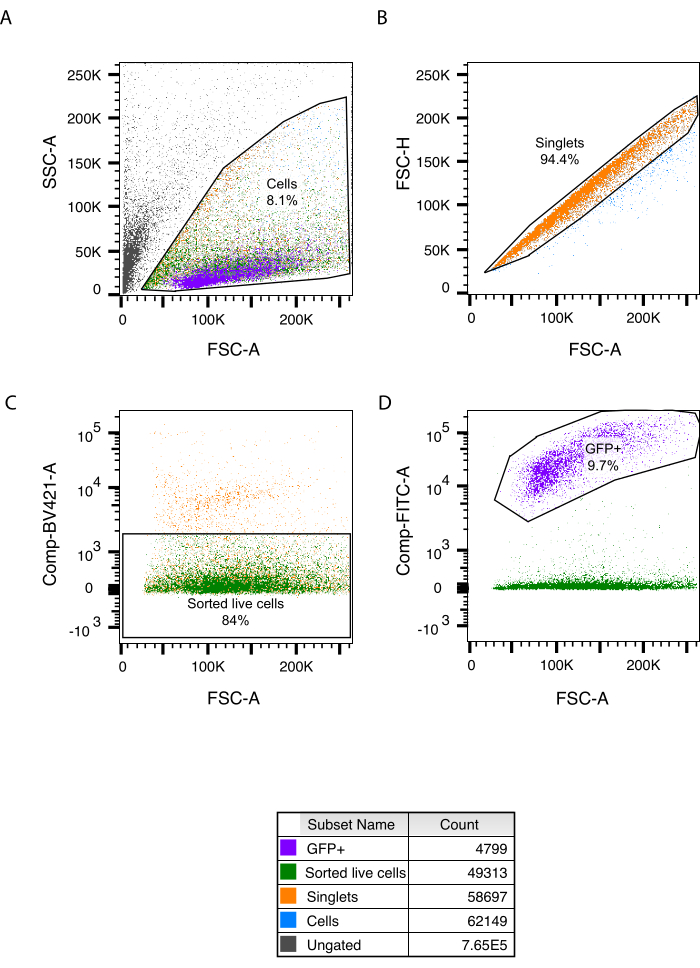
Figure 2: Gating strategy to sort live, single cells from the zebrafish gut. (A) Post sort analysis with FCS/SSC size gating. (B) Doublet discrimination. (C) Live/dead cell gating and sorted population. (D) Phox2bb:GFP+ cells present in the sorted live population. In each panel, the percentage of gated cells is shown. Abbreviations: FCS-A = forward scatter-peak area; SSC-A = side scatter-peak area; FSC-H = forward scatter-peak height; GFP = green fluorescent protein; FITC = fluorescein isothiocyanate. Please click here to view a larger version of this figure.
Supplemental Figure S1: Gut isolation and gating strategy to sort live, single cells from whole wildtype zebrafish larvae. (A) Brightfield image of gut isolation from 5 days post fertilization zebrafish larvae. The red line indicates the dissection line. (B) Post-sort analysis, showing all cells with FCS/SSC size gating. (C) Singlet selection. (D) DAPI gating. (E) Phox2bb:GFP+ gating. The percentage of gated cells is shown in each panel. Abbreviations: FCS-A = forward scatter-peak area; SSC-A = side scatter-peak area; FSC-H = forward scatter-peak height; GFP = green fluorescent protein; FITC = fluorescein isothiocyanate. Please click here to download this File.
