Optimization of ionomycin concentration
While ionomycin is required to induce eryptosis, increased ionomycin concentrations can lead to hemolysis (i.e. lysis of erythrocytes and release of hemoglobin), which needs to be avoided. Treatment of erythrocytes with 1 µM ionomycin in Ringer solution for 2 h is enough to induce eryptosis, as evidenced by successful labeling with annexin-V Alexa Flour 488 conjugate and quantification by FACS analysis (Figure 1A). Higher concentrations of ionomycin (5 and 10 µM) result in a slight increase in eryptosis (Figure 1A-D). However, such concentrations also enhance hemolysis (Figure 1E), which is not desired. In order to stay below 5% hemolysis, 1 µM ionomycin should be used.
Treatment time with ionomycin
Incubation of erythrocytes with ionomycin in Ringer solution for as little as 30 min is enough to induce eryptosis (Figure 2A). Increased incubation time increases the level of eryptosis, as measured by the annexin V-binding assay, for up to 2 h (Figure 2B,C). However, further incubation time results in a slight decrease in the level of eryptosis (Figure 2D). Maximum eryptosis was obtained after 2 h of treatment with 1 µM ionomycin, and for all other treatment times, lower eryptosis was obtained (Figure 2E). Representative flow cytometry histograms are presented in Figure 2A-D. In addition, average percentage eryptosis and hemolysis, for various treatment times with 1 µM ionomycin, are presented in Figure 2E and Figure 2F, respectively. The higher value of hemolysis after 180 min explains the reduction in eryptosis after the same amount of incubation (Figure 2E) as less viable cells exist upon 180 min of treatment with ionomycin.
Moreover, cells were treated with low concentrations of ionomycin including 0, 0.25, 0.5, and 1 µM for longer treatment times including 6 and 12 h, and eryptosis was measured (Figure 3). Cells treated with ionomycin concentrations of lower that 1 µM for 6 and 12 h show lower eryptosis compared to the cells treated with 1 µM ionomycin (Figure 3). Since decreasing the concentration and increasing the exposure time did not enhance eryptosis, 1 µM was used to trigger eryptosis.
Eryptosis is dependent on incubation time and extracellular glucose concentration
Extracellular glucose concentration affects the outcome of the process. Higher eryptosis values are observed when erythrocytes are pre-incubated in glucose-free Ringer solution compared to glucose-containing Ringer solution prior to incubation with 1 µM ionomycin for 2 h. The highest eryptosis values are obtained after 7 days of pre-incubation in both solutions. However, eryptosis is higher after pre-incubation in glucose-free Ringer solution compared to normal Ringer solution, which contains 5 mM glucose (see Figure 4A for representative plots and Figure 4B for comparison of global means). In addition, forward scatter histograms indicate the effect of glucose depletion on erythrocyte shrinkage (Figure 5A-D). Forward scatter is a measure for cell size based on the light refraction, and the level of light scattered is directly proportional to the size of cells33. The cells incubated in glucose-free Ringer solution show less forward scatter compared to the cells incubated in glucose-containing buffer (Figure 5E), indicating cell shrinkage in the glucose-free environment.
In addition to flow cytometry measurements, cells were observed under a confocal fluorescence microscope to confirm eryptosis. Erythrocytes with no treatment (Figure 6A) and with ionomycin treatment (Figure 6B) were labeled with annexin-V Alexa Flour 488 conjugate and observed under microscope. Treated cells showed a bright fluorescence signal (Figure 6B) due to the binding of annexin-V to PS in the outer leaflet. In contrast, cells with no treatment showed a very weak fluorescence signal (Figure 6A) indicating very low eryptosis. Further example images of eryptotic erythrocytes labeled with annexin-V with high fluorescence signal are shown in Figure 6C.
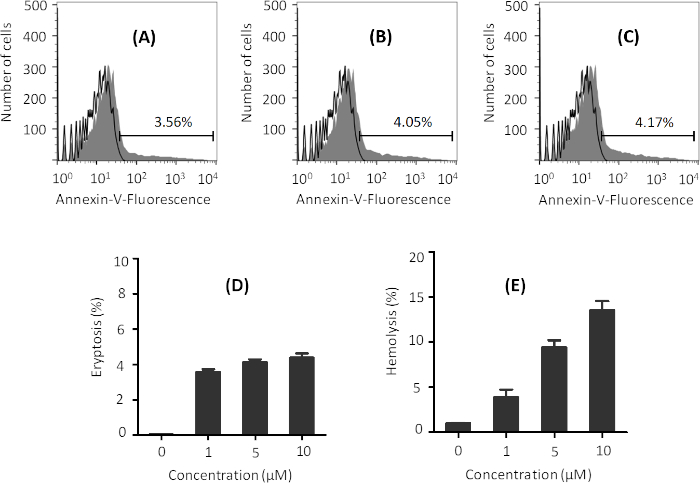
Figure 1: Representative graphs of the effect of various ionomycin concentrations on eryptosis and hemolysis. Flow cytometry histograms of erythrocytes treated with (A) 1 µM, (B) 5 µM, and (C) 10 µM ionomycin (gray) at 37 °C at 0.4% hematocrit in Ringer solution for 2 h. Black line indicates non-treated cells. Percentage of eryptosis is indicated in each figure. Phosphatidylserine exposure was measured using annexin-V binding. (D) Arithmetic means ± SD (n = 3) of the percentage eryptosis of cells treated with different concentrations of ionomycin after 2 h treatment, and (E) arithmetic means ± SD (n = 3) of hemolysis of erythrocytes by different concentrations of ionomycin under same conditions. Please click here to view a larger version of this figure.
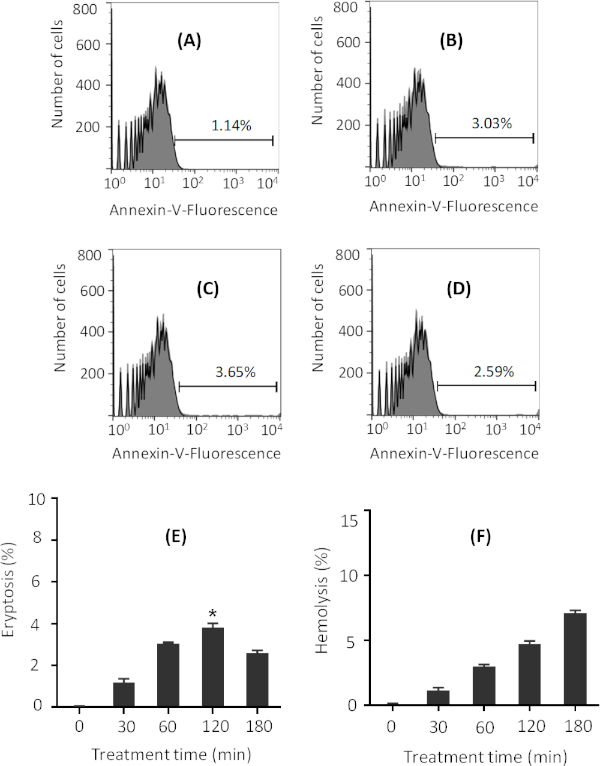
Figure 2: Representative figures on the effect of various ionomycin treatment times on eryptosis. Flow cytometry histograms of erythrocytes treated with 1 µM ionomycin (gray) at 37 °C for (A) 30 min, (B) 60 min, (C) 120 min, and (D) 180 min at 0.4% hematocrit in Ringer solution. Black line indicates non-treated cells. Percentage of eryptosis is indicated in each figure. Phosphatidylserine exposure was measured through annexin-V binding. (E) Arithmetic means ± SD (n = 3) of percentage eryptosis of cells treated with 1 µM ionomycin for different times. The highest eryptosis was obtained after 120 min treatment. (F) Arithmetic means ± SD (n = 3) of percentage hemolysis of cells treated with 1 µM ionomycin for different times. For statistical analysis, one-way non-parametric ANOVA with Kruskal-Wallis test was performed, and eryptosis after 120 min treatment was significantly higher than control as indicated in panel E. * is for p < 0.05. Please click here to view a larger version of this figure.
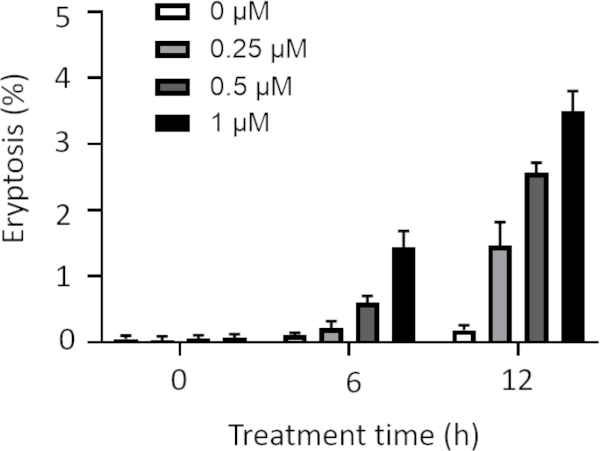
Figure 3: Effect of various ionomycin concentrations and treatment times on eryptosis. Arithmetic means ± SD (n = 3) of the percentage eryptosis of cells treated with different concentrations of ionomycin is shown after various treatment times. The cells were treated with low concentrations of ionomycin including 0, 0.25, 0.5, and 1 µM for longer exposure (6 h and 12 h). Higher concentrations and longer treatments resulted in higher eryptosis values. Please click here to view a larger version of this figure.
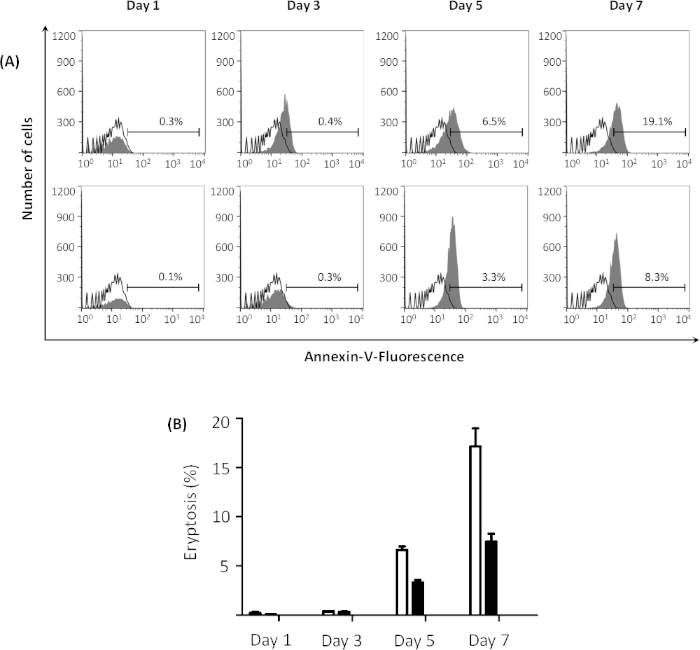
Figure 4: Effect of energy depletion on eryptosis. (A) Flow cytometry histogram for erythrocytes treated with 1 µM ionomycin (gray) at 37 °C for 2 h at 0.4% hematocrit, after pre-incubation in glucose-free Ringer solution (top figures) and Ringer solution (bottom figures) from 1 to 7 days, reveals that energy depletion facilitates eryptosis. Black line indicates non-treated cells. Percentages of eryptosis are indicated in the graphs for each day. (B) Arithmetic means ± SD (n = 3) of the percentage eryptosis of erythrocytes treated with 1 µM ionomycin at 37 °C for 2 h at 0.4% hematocrit, after pre-incubation in Ringer solution (black bars) and glucose-free Ringer solution (white bars) from 1 to 7 days. Please click here to view a larger version of this figure.
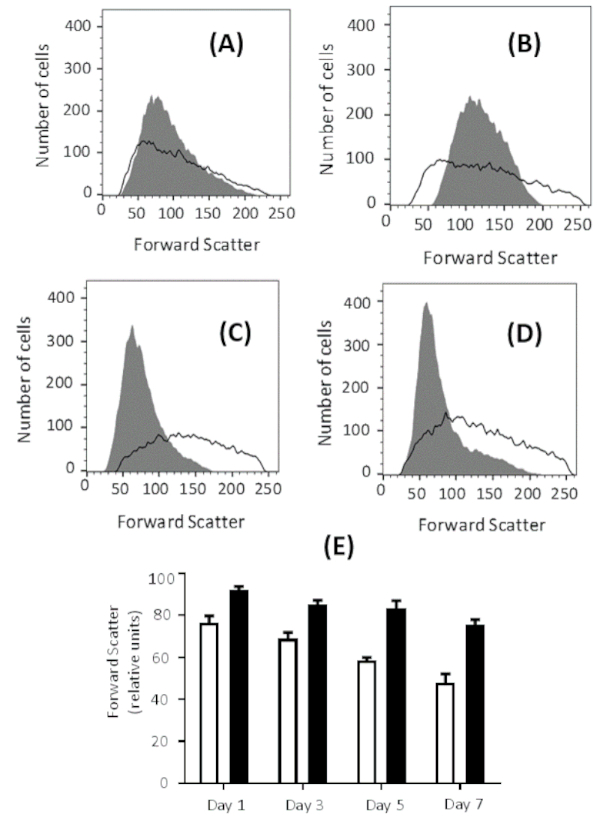
Figure 5: Effect of energy depletion on cell size. Forward scatter histogram for erythrocytes treated with 1 µM ionomycin at 37 °C for 2 h at 0.4% hematocrit, after pre-incubation in glucose-free Ringer solution (gray) and Ringer solution (black line) for (A) 1 day, (B) 3 days, (C) 5 days, and (D) 7 days. The forward scatter histogram over time indicates erythrocyte shrinkage in glucose-free buffer. (E) Arithmetic means ± SD (n = 3) of forward scatter intensities of erythrocytes treated with 1 µM ionomycin at 37 °C for 2 h at 0.4% hematocrit, after pre-incubation in Ringer solution (black bars) and glucose-free Ringer solution (white bars) from 1 to 7 days. Please click here to view a larger version of this figure.

Figure 6: Confocal fluorescence microscopy images of erythrocytes treated with (A) 0 µM, (B) and (C) 1 µM ionomycin at 37 °C for 2 h at 0.4% hematocrit. 40x objective magnification was used for images in panels A and B, and 100x objective magnification was used to take images for panel C. PS in healthy erythrocytes is located on the inner leaflet of the cell membrane, therefore there is no fluorescence signal in panel A. In panels B and C erythrocytes have been induced for eryptosis and there is a bright fluorescence signal resulting from the binding of annexin-V to PS translocated to the outer leaflet of the cell membrane. Please click here to view a larger version of this figure.
| Cell density /hematocrit | Ionomycin concentration | Buffer | Pre-incubation | Treatment time with ionomycin | Detection method | Reference |
| 1.65 x 108 cells/mL | 0.3 mM | Buffer A* | 36 h in buffer A | 1 h | Annexin V | 12 |
| 0.40% | 1 mM | Ringer solution | 48 h in Ringer | 1 h | Annexin V | 17 |
| 50% | 10 mM | Buffer B** | – | 3 h | Merocyanine 540 | 18 |
| 0.40% | 1 mM | Ringer solution | 48 h in Ringer | 1 h | Annexin V | 19 |
| 0.40% | 1 mM | Ringer solution | 48 h in Ringer | 1 h | Annexin V | 20 |
| 2% | 1 mM | Ringer solution | – | 4 h | Annexin V | 21 |
| 0.40% | 1 mM | Ringer solution | – | 0.5 h | Annexin V | 22 |
| 10% | 1 mM | Ringer solution | – | 3 h | Annexin V | 23 |
| 0.40% | 10 mM | Ringer solution | – | 0.5 h | Annexin V | 24 |
| 0.40% | 1 mM | Ringer solution | 48 h in Ringer | 0.5 h | Annexin V | 25 |
| 2 x 106 cells/mL | 1 mM | HEPES-buffered saline (HBS) | – | 0.5 h | Annexin V | 26 |
| *Buffer A: 10 mM HEPES, 150 mM NaCl, 5 mM KCl, 1 mM MgCl2·6H2O, 10 mM glucose, and 1.8 mM CaCl2·2H2O | ||||||
| **Buffer B: 5 mM Tris, 100 mM KC1, 60 mM NaCl, and 10 mM glucose | ||||||
Table 1: Various protocols used in the literature to induce eryptosis using ionomycin.
