A murine IL-22 reporter transgene was created using recombineering to modify a bacterial artificial chromosome carrying the IL-22 locus. Figure 1 shows a diagram of pBACe3.6 vector containing the sacBII gene, a positive-selection marker, and chloramphenicol antibiotic resistance gene11. After introducing tdTomato into exon 1, the signal peptide sequence was disrupted, as shown in Figure 2. Thus, the tdTomato reporter was trapped inside the IL-22-expressing cells, enabling their detection and isolation by flow cytometry. The homozygous mice were bred from founder lines and screened by PCR, and they did not require backcrossing in order to achieve offspring with a genetic identity, as is required in the usual knock-in strategy. To test the fidelity of the IL-22 reporters, in vitro-generated splenocytes were cultured under Th22 or neutral conditions. Figure 3 demonstrates that IL-22-tdtomato was well expressed in Th22 cells in vitro, detected either by flow cytometry or by fluorescent microscopy. In vivo, as shown in Figure 4, the IL-22 reporter was detected in different mouse tissues under homeostatic condition. Most of the tdTomato signal was found in the lamina propria (LP) cells from the gut, but not in the axillary lymph node (ALN), the spleen, or the thymus. To visualize the tdTomato reporter in inflamed tissue, we used a mouse colitis model induced by the transfer of reporter CD4+CD45Rbhi T cells into Rag1-/- mice. Figure 5 demonstrates that the reporters were first present in the mesenteric lymph nodes and then accumulated inside the lamina propria of distal small intestine and colon tissues. Taken together, this novel method for the generation of IL-22 reporter mice is effective and time-saving.
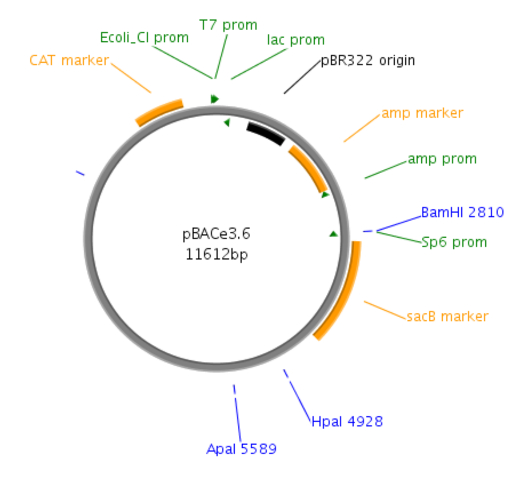
Figure 1: Site map of the pBACe3.6 vector. pBACe3.6 is characterized by containing chloramphenicol antibiotic resistance and a sacBII gene as a positive-selection marker. During recombination, the desired clones are sucrose-resistant through their expressing the sacBII gene and are allowed to grow on sucrose-containing media, while the negative BAC colonies are sucrose-sensitive.
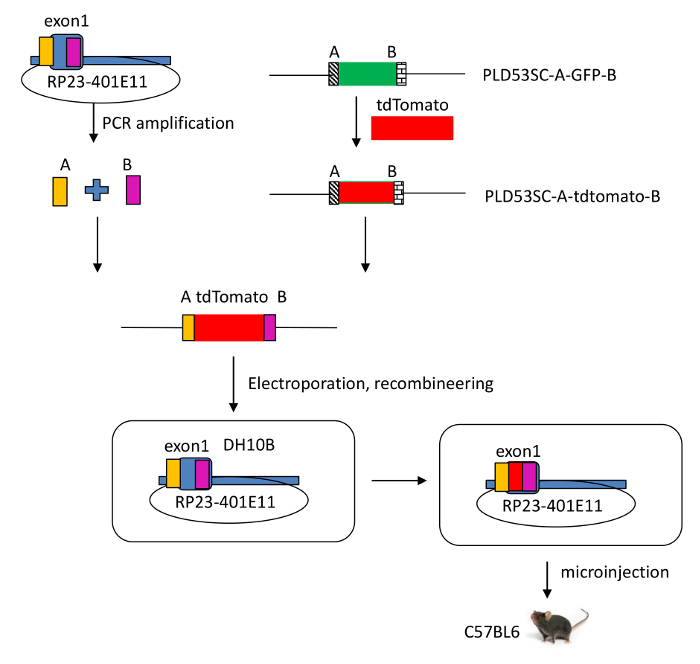
Figure 2: Schematic view of the modified RP23-401E11 BAC clone. A tdTomato reporter gene was introduced into the Il-22 locus, which includes IL-22 and regulatory elements in a murine bacterial artificial chromosome (BAC RP23-401E11), using recombineering technology. By homologous recombination, the sequence of the signal peptide of Il-22 in the BAC was disrupted and exon 1 of Il-22 was replaced by the tdTomato reporter gene. Please click here to view a larger version of this figure.
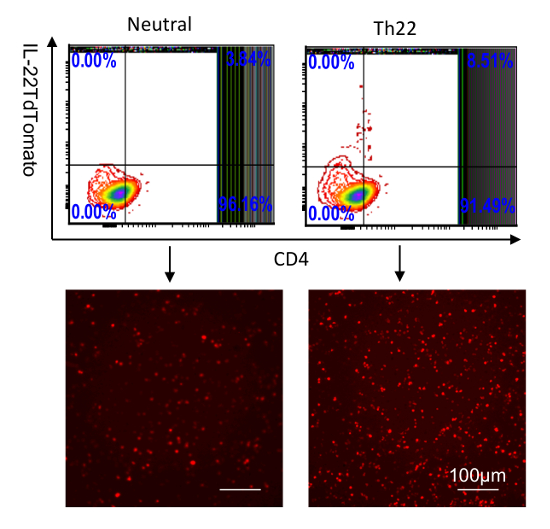
Figure 3: Identification of IL-22 reporters in splenocytes cultured in vitro. CD4 T cells were purified from the spleen and then stimulated with anti-CD28; anti-CD3 (neutral condition); or anti-CD3, anti-CD28, anti-IFNƳ, anti-IL4, IL-6, TGF-β, and 6-Formylindolo (3,2-b)carbazole (FICZ) for 5 days. IL-22-tdTomato was analyzed by flow cytometry (top two) and fluorescent microscopy (bottom two, scale bar: 100 µm). The numbers in the quadrants indicate the percent of CD45+ cells. Please click here to view a larger version of this figure.
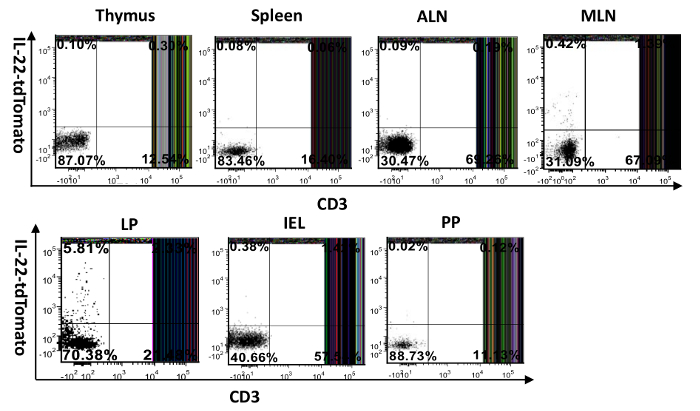
Figure 4: Visualization of IL-22-producing cells in different mouse tissues. Single-cell suspensions were prepared from the spleen, thymus, lymph nodes, intestines (IEL and LP), and Peyer's patch. They were then surface-stained with FITC-anti-CD3 and examined for tdTomato expression. MLN: mesenteric lymph node, ALN: axillary lymph node, PP: Peyer's patch, IEL: intraepithelial cells isolated from the small intestine, LP: lamina propria cells purified from the small intestine. The numbers in the quadrants indicate the percent of CD45+ cells. Please click here to view a larger version of this figure.
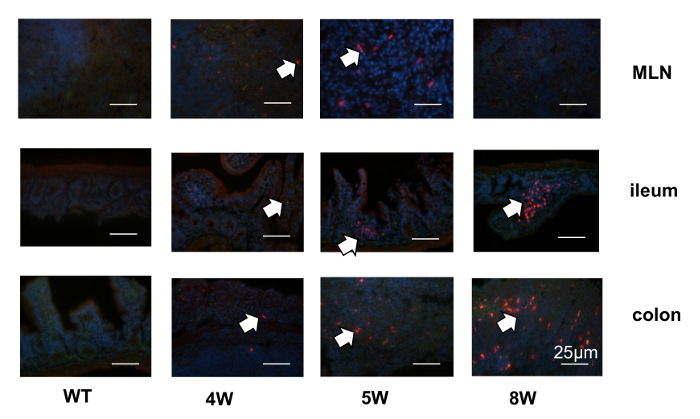
Figure 5: Localization of IL-22 reporters during the development of T-cell transfer colitis. CD4CD45RBHigh T cells from IL-22 reporter mice were transferred into Rag1-/- mice, and the tdTomato signals were evaluated by immunohistochemistry in the different tissues at different time points during the development of colitis. The arrows point to the IL-22-tdTomato signals. Scale bars = 25 µm. Please click here to view a larger version of this figure.
