Pollination studies in apricot require the use of flowers at the late balloon stage one day before anthesis (Figure 1A). This stage is considered the most favorable for both pollen and pistil collection, since floral structures are nearly mature, but anther dehiscence has not yet occurred. This prevents the interference of undesired pollen, not only of pollen from the same flower but also from other flowers, since the closed petals impede the arrival of insects carrying external pollen. The pollen grains are easily sieved through a fine mesh (Figure 1B) from dehisced anthers previously placed on a piece of paper for 24 h at room temperature or with slight extra heat. Likewise, pistils are obtained from flowers at balloon stage after the removing of petals, sepals and stamens with the help of tweezers or fingernails (Figure 1C). Pistils can be self- and cross-pollinated with a fine brush (Figure 1D).
The hermaphroditic flowers of apricot have five dark red sepals, five white petals (Figure 1A), a single pistil (Figure 2A) and 25-30 stamens. The pistil has three main structures: stigma, style and ovary. The ovary has two ovules, and the fertilization of at least one of them is required for fruit setting. During pollination, insects, mainly bees, transfer pollen grains to the stigma (Figure 1A), where they germinate (Figure 2B) within 24 h following pollination. A pollen tube is produced from each germinating pollen grain, which grows through the pistil structures to reach the ovary after 3-4 days and fertilize one of the two ovules after around 7 days. In self-incompatible cultivars in which the S allele of the pollen grain is the same as one of the two of the pistils, pollen tube stops growing at the upper style, preventing fertilization (Figure 2C). However, the pollen tubes from a compatible cultivar, with a different S allele, can grow through the style (Figure 2D), reach the ovary (Figure 2E) and fertilize one of the two ovules.
The analysis of in vitro pollen germination showed good pollen viability in all the cultivars analyzed here, since most pollen tubes were longer than the length of the pollen grain after 24 h in the culture medium. Germinated pollen grains were observed at the stigma surface (Figure 2B) in pistils from all pollinations, indicating adequate pollination (Figure 3).
To determine the self-(in)compatibility for each cultivar, pollen tube behavior in self- and cross-pollinations done in laboratory-controlled conditions was observed under fluorescence microscopy. Pollen tube growth was recorded along the style in all the pistils examined. Cultivars were considered as self-incompatible when pollen tube growth was arrested along the style in most self-pollinated pistils (Figure 2C, Figure 3) and self-compatible when at least one pollen tube reached the base of the style in most of the pistils examined (Figure 2E, Figure 3).
The study of the S-locus by PCR analysis allowed characterizing the S-genotype of each cultivar. Firstly, the S-alleles were identified by the amplification of the first S-RNase intron using the primers SRc-F/SRc-R (Table 2). The size of the amplified fragments was analyzed by capillary electrophoresis (Figure 4A) and was used to classify the genotypes analyzed in their corresponding incompatibility group (I.G.) (Table 3).
Some pairs of alleles, such as S1 and S7 or S6 and S9, showed similar fragment sizes for the first intron. Thus, the differentiation of these alleles was done by amplifying a region of the second intron of the RNase with the primers Pru-C2/PruC4R, SHLM1/SHLM2 and SHLM3/SHLM4 (Table 2). The PruC2/PruC4R primer combination was used to distinguish between S6 and S9. For S6, a fragment of 1300 bp was amplified whereas a fragment of around 700 bp was observed for the S9 allele (Figure 4B, Table 3). The specific primers SHLM1/SHLM2 and SHLM3/SHLM4 amplified a fragment of approximately 650 bp in the S1 allele and 413 bp in the S7 allele (Figure 4C, Table 3).
The primers AprFBC8-(F/R) that amplify the V2 and HVb variable regions of the SFB gene were used to distinguish Sc and S8 alleles since both alleles show identical RNase sequence. The S8 allele showed a PCR-fragment of approximately 150 bp whereas a 500 bp fragment corresponded to the Sc allele (Figure 4D, Table 3). Once the S-genotype was determined for all the cultivars, self-incompatible cultivars were assigned to their corresponding incompatibility groups based on their S-alleles (Table 3).
This approach requires determining the self-(in)compatibility of each cultivar by controlled self- and cross-pollinations in the laboratory (Figure 5A) concomitantly with the characterization of the S-genotype by genetic analysis (Figure 5B). As a result, the pollination requirements of each cultivar and the incompatibility relationships among apricot cultivars can be determined.

Figure 1. Experimental set up for the determination of self-(in)compatibility in apricot.
(A) Flowers at balloon stage (black arrows) in the field. (B) Sieve of pollen grains using a fine mesh. (C) Pistils placed on florist foam in water. (D) Hand-pollination of the pistils with the help of a paintbrush. Please click here to view a larger version of this figure.
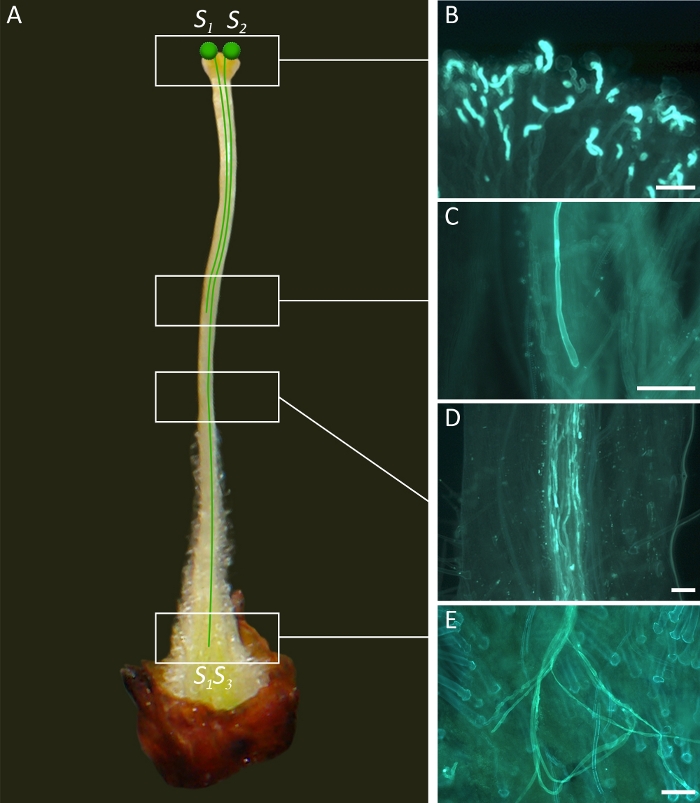
Figure 2. Diagrammatic representation of gametophytic incompatibility relationships in apricot flowers.
(A) In Gametophytic Self-Incompatibility (GSI), both compatible and incompatible pollen grains germinate on the stigma. The pollen grain carries one of two S-alleles of the original genotype, in this case either S1 or S2. If the S-allele of the pollen grain matches one of the two S-alleles of the pistil, in this case S1S3, pollen tube growth is inhibited in the upper one-third of the style. (B) Germination of pollen grains on the stigma surface. (C) Pollen tube arrested in the style indicating an incompatible behavior. (D) Pollen tubes growing along the style. (E) Pollen tubes at the base of the style indicating a compatible behavior. Scale bars, 100 μm. Please click here to view a larger version of this figure.
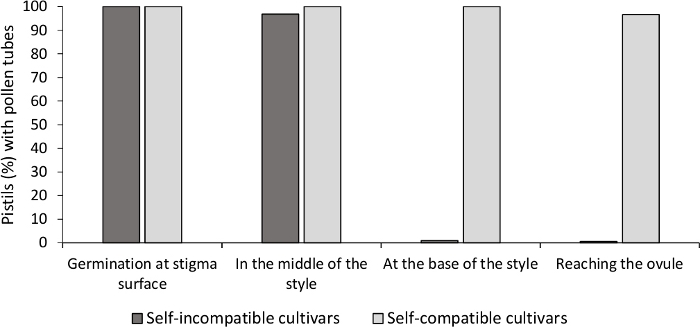
Figure 3. Representative results of pollen germination and pollen tube growth through the style for self-compatible and self-incompatible cultivars after self-pollinations.
Percentage of pistils with pollen grains germinating at the stigma surface, with pollen tubes in halfway the style, at the base of the style, and reaching the ovule. Please click here to view a larger version of this figure.
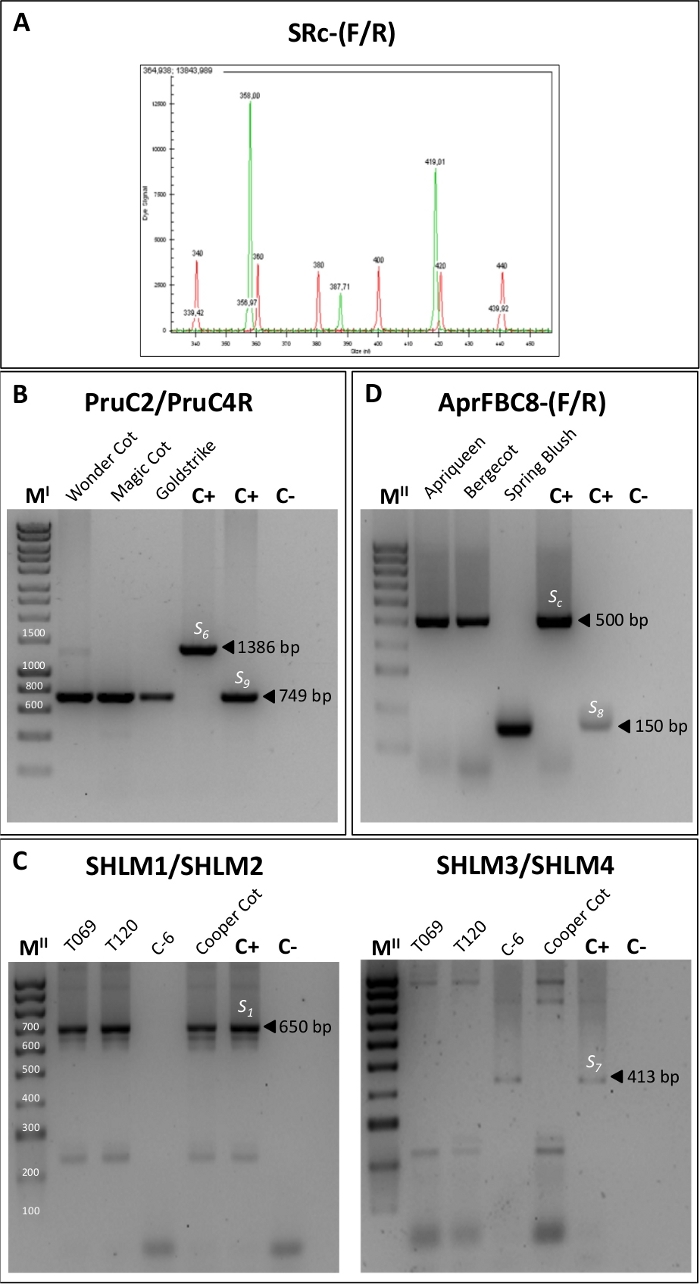
Figure 4. PCR fragment amplification using five primer pair combinations for the identification of S-alleles.
(A) Gene analyzer output for the SRc-(F/R) primers showing the size of the two amplified fragments of the RNase first intron region corresponding to the S-alleles. (B) PCR amplification using the primers PruC2/PruC4R for the identification of the S6 and S9 alleles. (C) PCR products obtained using the specific primers SHLM1 and SHLM2 for the differentiation of the S1 allele and SHLM3 and SHLM4 to distinguish the S7 allele. (D) PCR amplification with the AprFBC8-(F/R) primers for identifying Sc and S8 alleles. MI: 1 kb DNA Ladder. MII: 100 bp DNA Ladder. Please click here to view a larger version of this figure.
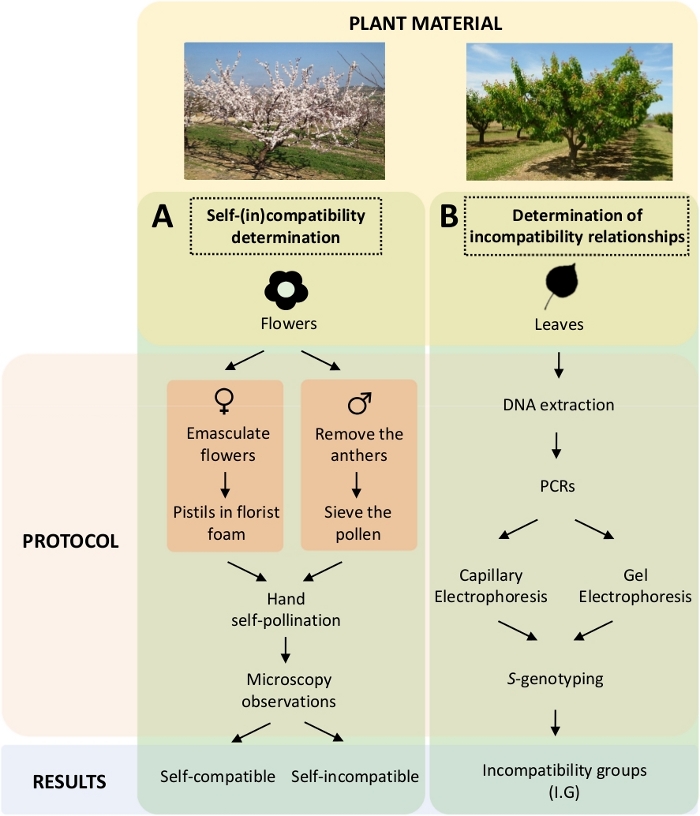
Figure 5. Scheme of the experimental design to elucidate the self- and inter-(in) compatibility relationships in apricot cultivars.
(A) Workflow of self-(in)compatibility determination by controlled pollinations in the laboratory. (B) Workflow of the S-allele identification by molecular approaches. Please click here to view a larger version of this figure.
| PCR Master Mix | Thermocycler conditions | ||||||
| Components | Final Concentration | 15 μL reaction | Cycle Step | Temperature | Time | Cycles | |
| 10x NH4 Reaction Buffer | 10x | 1.5 μL | Initial denaturation | 94 °C | 3 min | 1 | |
| 50 mM MgCl2 Solution | 25 mM | 1.2 μL | Denaturing | 94 °C | 1 min | 35 | |
| 100 mM dNTP | 2.5 mM | 0.6 μL | Annealing | 55 °C | 1 min | ||
| Primer SRc-F | 10 μM | 0.6 μL | Extension | 72 °C | 3 min | ||
| Primer SRc-R | 10 μM | 0.6 μL | Final Extension | 72 °C | 5 min | 1 | |
| 500 U Taq DNA Polymerase | 0.5 U | 0.2 μL | 4 °C | hold | |||
| H2O | 8.3 μL | ||||||
| Components | Final Concentration | 25 μL reaction | Cycle Step | Temperature | Time | Cycles | |
| 10x PCR buffer | 10x | 2.5 μL | Initial denaturation | 94 °C | 2 min | 1 | |
| 5x Q-solution | 5x | 5 μL | Denaturing | 94 °C | 10 s | 10 | |
| 100 mM dNTP | 2.5 mM | 0.5 μL | Annealing | 55 °C | 2 min | ||
| Primer PruC2 | 10 μM | 0.2 μL | Extension | 68 °C | 2 min | ||
| Primer C4R | 10 μM | 0.2 μL | Denaturing | 94 °C | 10 s | 25 | |
| 250 U Taq DNA Polymerase | 10 U | 0.13 μL | Annealing | 58 °C | 2 min | ||
| H2O | 15.5 μL | Extension* | 68 °C | 2 min | |||
| Final Extension | 72 °C | 5 min | 1 | ||||
| 4 °C | hold | ||||||
| * with 10 s added each cycle to the 68 %C extension step. | |||||||
| Components | Final Concentration | 25 μL reaction | Cycle Step | Temperature | Time | Cycles | |
| 10x PCR buffer | 10x | 2.5 μL | Initial denaturation | 94 °C | 2 min | 1 | |
| 5x Q-solution | 5x | 5 μL | Denaturing | 94 °C | 30 s | 35 | |
| 100 mM dNTP | 2.5 mM | 0.5 μL | Annealing | 62 °C | 1.5 min | ||
| Primer SHLM1 | 10 μM | 0.2 μL | Extension | 72 °C | 2 min | ||
| Primer SHLM2 | 10 μM | 0.2 μL | Final Extension | 72 °C | 5 min | 1 | |
| 250 U Taq DNA Polymerase | 10 U | 0.13 μL | 4 °C | hold | |||
| H2O | 15.5 μL | ||||||
| Components | Final Concentration | 20 μL reaction | Cycle Step | Temperature | Time | Cycles | |
| 5x PCR Buffer | 5x | 4 μL | Initial denaturation | 98 °C | 30 s | 1 | |
| dNTP | 2.5 mM | 1.6 μL | Denaturing | 98 °C | 10 s | 35 | |
| Primer SHLM3 | 10 μM | 1 μL | Annealing | 51 °C | 30 s | ||
| Primer SHLM4 | 10 μM | 1 μL | Extension | 72 °C | 1 min | ||
| 100 U DNA Polymerase | 5 U | 0.2 μL | Final Extension | 72 °C | 5 min | 1 | |
| H2O | 12.4 μL | 4 °C | hold | ||||
| Components | Final Concentration | 25 μL reaction | Cycle Step | Temperature | Time | Cycles | |
| 10x PCR buffer | 10x | 2.5 μL | Initial denaturation | 94 °C | 2 min | 1 | |
| 100 mM dNTP | 2.5 mM | 2 μL | Denaturing | 94 °C | 30 s | 35 | |
| Primer FBC8-F | 10 μM | 1 μL | Annealing | 55 °C | 1.5 min | ||
| Primer FBC8-R | 10 μM | 1 μL | Extension | 72 °C | 2 min | ||
| 250 U Taq DNA Polymerase | 10 U | 0.125 μL | Final Extension | 72 °C | 5 min | 1 | |
| H2O | 17.4 μL | 4 °C | hold | ||||
Table 1. Reaction and cycling conditions for different primer combinations used in this protocol.
| Primers | Sequence | Reference |
| SRc-F | 5'-CTCGCTTTCCTTGTTCTTGC-3' | 18 |
| SRc-R | 5'-GGCCATTGTTGCACCCCTTG-3' | 18 |
| Pru-C2 | 5'-CTTTGGCCAAGTAATTATTCAAACC-3' | 35 |
| Pru-C4R | 5'-GGATGTGGTACGATTGAAGCG-3' | 35 |
| SHLM1-F | 5'-GGTGGAGGTGATAAGGTAGCC-3' | 17 |
| SHLM2-R | 5'-GGCTGCATAAGGAAGCTGTAGG-3' | 17 |
| SHLM3-F | 5'-TATATCTTACTCTTTGGC-3' | 17 |
| SHLM4-R | 5'-CACTATGATAATGTGTATG-3' | 17 |
| AprFBC8-F | 5'-CATGGAAAAAGCTGACTTATGG-3' | 26 |
| AprFBC8-R | 5'-GCCTCTAATGTCATCTACTCTTAG-3' | 26 |
Table 2. Primers used in this protocol, sequence and reference for S-genotype characterization in Prunus armeniaca.
| Cultivar | SRc-(F/R) (bp) | PruC2/PruC4R (bp) | SHLM1/SHLM2 (bp) | SHLM3/SHLM4 (bp) | AprFBC8-(F/R) (bp) | S-Genotype | Incompatibility group (I.G) |
| Wonder Cot8 | 420, 420 | 749, 1386 | S6S9 | VIII | |||
| Magic Cot8 | 334, 420 | 749 | S2S9 | XX | |||
| Goldstrike8 | 334, 420 | 749 | S2S9 | – | |||
| T06917 | 334, 408 | 650 | S1S2 | I | |||
| T12017 | 334, 408 | 650 | S1S2 | – | |||
| C-6 | 334, 408 | 413 | S2S7 | IV | |||
| Cooper Cot8 | 274, 408 | 650 | S1S3 | XVIII | |||
| Apriqueen | 358, 358 | 500 | ScSc | – | |||
| Bergecot8 | 334, 358 | 500 | S2Sc | – | |||
| Spring Blush8 | 274, 358 | 150 | S3S8 | XXI |
Table 3. S-genotyping of apricot cultivars with five primer pairs used in this protocol and incompatibility group assignment. The different polymerase chain reaction product sizes of S-alleles amplified using SRc-(F/R), PruC2/PruC4R, SHLM1/ SHLM2, SHLM3/SHLM4, and AprFBC8-(F/R) primers are shown in the table.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}