1. Buffers and stock solutions
- Make a 1 M 3-morpholinopropane-1-sulfonic acid (MOPS) solution in deionized water, pH 7.4. Store at 4 °C.
- Prepare 500 mM ethylenediaminetetraacetic acid (EDTA) in deionized water, pH 8.0. Store at room temperature.
- Prepare 2.4 M sorbitol in deionized water. Store at room temperature after autoclaving.
- Prepare 2.5 M sucrose in deionized water. Store at room temperature after autoclaving.
- Prepare 200 mM phenylmethylsulfonyl fluoride (PMSF) in isopropanol. Store at −20 °C.
CAUTION: PMSF is toxic if swallowed and causes severe skin burns and eye damage. Do not breathe in the dust, and wear protective gloves and goggles. - Prepare 72% trichloroacetic acid (TCA) in deionized water. Store at 4 °C.
CAUTION: TCA may cause respiratory irritation, severe skin burns, and eye damage. Do not breathe in the dust, and wear protective gloves and goggles. - Prepare the SM buffer: 20 mM MOPS and 0.6 M sorbitol, pH 7.4.
- Prepare the swelling buffer: 20 mM MOPS, 0.5 mM EDTA, 1 mM PMSF, and protease inhibitor cocktail, pH 7.4.
- Prepare the sucrose gradient buffers: 0.8 M, 0.96 M, 1.02 M, 1.13 M, and 1.3 M sucrose in 20 mM MOPS, and 0.5 mM EDTA, pH 7.4.
NOTE: All the buffers can be stored at 4 °C; however, it is recommended to store the sucrose-containing buffers at −20 °C to avoid fungal growth. The protease inhibitors have to be added freshly prior to use.
2. Generation of submitochondrial vesicles
- Isolate crude yeast mitochondria freshly according to established protocols34,35.
NOTE: For this experiment, mitochondria were isolated from Saccharomyces cerevisiae. The generation of gradient pure mitochondria devoid of other organelles is not necessary. - Resuspend 10 mg of freshly isolated mitochondria in 1.6 mL SM buffer (4 °C) by pipetting (Figure 2, step 1).
- Transfer the mitochondrial suspension to a pre-cooled 100 mL Erlenmeyer flask.
- Slowly add 16 mL of swelling buffer using a 20 mL glass pipette. Apply constant mild stirring on ice during the addition. This osmotic treatment results in water uptake into the matrix space. The swelling of the mitochondrial inner membrane will disrupt the outer membrane. However, both membranes will stay in contact at the contact sites9(Figure 2, step 2).
- Incubate the samples under constant mild stirring for 30 min on ice.
- Slowly add 5 mL of 2.5 M sucrose solution using a 5 mL glass pipette to increase the sucrose concentration in the samples to approximately 0.55 M. This osmotic treatment results in a water efflux from the matrix36. The shrinkage of the inner membrane is intended to maximize its distance to the residual fragments of the outer membrane. This decreases the probability of generating artificial hybrid vesicles of inner and outer membranes through sonication (Figure 2, step 3).
- Incubate the samples under mild stirring for 15 min on ice.
- Subject the mitochondria to sonication to generate submitochondrial membrane vesicles (Figure 2, step 4).
- Transfer the mitochondrial suspension to a pre-cooled rosette cell.
- Sonicate the mitochondrial suspension at a 10% amplitude for 30 s while cooling the rosette cell in an ice bath.
- Rest the suspension for 30 s in an ice bath.
- Repeat step 2.8.2 and step 2.8.3 three more times.
NOTE: The sonication conditions will vary according to the sonicator used. Optimization will be necessary for individual machines. Sonication that is too harsh will lead to the artificial formation of vesicles consisting of mitochondrial inner and outer membranes.
3. Separation of submitochondrial vesicles
- Separate the generated vesicles from the remaining intact mitochondria by centrifugation at 20,000 x g for 20 min at 4 °C. The intact mitochondria will pellet while the vesicles will stay in the supernatant.
- Concentrate the vesicle mixture.
- Transfer the supernatant to fresh ultracentrifugation tubes.
- Load a cushion of 0.3 mL of 2.5 M sucrose solution at the bottom of the tube using a 1 mL syringe equipped with a 0.8 mm x 120 mm cannula.
- Centrifuge at 118,000 x g for 100 min at 4 °C.
NOTE: Here, a swinging bucket rotor with a maximum radius of 153.1 mm was used. The centrifugation conditions strongly depend on the rotor and will have to be adapted when another rotor is used.
- The vesicles will appear as a disc on the top of the sucrose cushion after the centrifugation. Discard approximately 90% of the supernatant. Now, harvest the concentrated vesicles by resuspending them in the remaining buffer including the 2.5 M sucrose by pipetting up and down. Transfer the suspension to an ice-cold Dounce homogenizer.
- Homogenize the suspension with at least 10 strokes using a polytetrafluoroethylene potter.
- Prepare the sucrose gradient.
- For an 11 mL step gradient with five steps (1.3 M, 1.13 M, 1.02 M, 0.96 M, and 0.8 M sucrose in 20 mM MOPS and 0.5 mM EDTA, pH 7.4), each layer has 2.2 mL of sucrose solution.
NOTE: A refractometer is recommended to measure and adjust the sucrose concentrations; however, it is not essential. Apply 10 µL of the buffer to the refractometer, and detect the respective refractive indices. The calculation of sucrose concentration is as follows:
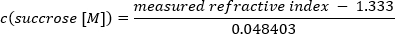
1.3333 represents the refractive index of water. 0.048403 is a sucrose specific conversion index obtained from the linear scaling of molar refractivity to sucrose concentration in aqueous solutions 37. - Add the highest sucrose concentration to the centrifugation tube, and put the tube at −20 °C. Wait until the layer is completely frozen before applying the next one. Proceed accordingly until the last layer is added, and store the gradients at −20 °C.
NOTE: Remember to transfer the frozen gradients to 4 °C when starting the experiment to allow the gradients to thaw. This will take approximately 3 h. For the centrifugation here, a swinging bucket rotor with a capacity of 13.2 mL was used. When a different rotor is used, the gradient step volumes must be adapted accordingly.
- For an 11 mL step gradient with five steps (1.3 M, 1.13 M, 1.02 M, 0.96 M, and 0.8 M sucrose in 20 mM MOPS and 0.5 mM EDTA, pH 7.4), each layer has 2.2 mL of sucrose solution.
- Measure the sucrose concentration of the samples using a refractometer. If there is not access to a refractometer, one could alternatively assume that the sucrose concentration is 2 M. This supposed sucrose concentration will then be the basis for the next step.
- Adjust the sucrose concentration to 0.6 M by the addition of an appropriate amount of 20 mM MOPS, 0.5 mM EDTA, 1 mM PMSF, and protease inhibitor cocktail, pH 7.4. If the sucrose concentration is estimated as 2 M instead of being measured, it is recommended to test whether the concentration is low enough after adjustment. Apply a small aliquot of the sample (ca. 50 µL) on top of 200 µL of 0.8 M sucrose in a test tube. The sample must stay on top of the 0.8 M sucrose.
- Load the samples (approximately 1 mL) on top of the sucrose gradient (keep 10% for later reference). Careful pipetting is important to avoid the disturbance of the gradient (Figure 2, step 5).
- Separate the vesicles by centrifugation at 200,000 x g and 4 °C for 12 h. If possible, set the centrifuge to slow acceleration and deceleration to avoid the disturbance of the gradient (Figure 2, step 6).
NOTE: Here, a swinging bucket rotor with a maximum radius of 153.1 mm was used. The centrifugation conditions strongly depend on the rotor and will have to be adapted when another rotor is used. - Harvest the gradient from top to bottom in 700 µL fractions using a 1 mL pipette. This will result in 17 fractions, which allows for a sufficient resolution.
4. Analysis of submitochondrial vesicles
- Concentrate the proteins by subjecting each fraction to two sequentially performed TCA precipitations38 to prepare the SDS-PAGE samples.
- Add 200 µL of 72% TCA to the individual fractions, and mix until the solution is homogeneous. Incubate the fractions for 30 min on ice, and pellet the precipitated proteins by centrifugation at 20,000 x g and 4 °C for 20 min. Discard the supernatant, add 500 µL of 28 % TCA solution, mix well, and repeat the centrifugation step.
- Wash the pellets with 1 mL of acetone (−20°C), and centrifuge for 10 min at 20,000 x g and 4 °C. Discard the supernatant, and let the pellets air dry. Resuspend the pellets in 60 µL of SDS sample buffer, and incubate the samples for 5 min at 95 °C.
NOTE: A large amount of sucrose will remain, particularly in the high-density fractions, after the first TCA precipitation. This will be removed through the additional TCA precipitation.
- Analyze 20 µL of each fraction by SDS-PAGE39 and immunoblotting40,41,42,43.
It is relatively easy to separate mitochondrial inner and outer membranes. However, the generation and separation of contact site-containing vesicles are much more difficult. In our opinion, two steps are critical and essential: the sonication conditions and the gradient used.
Usually, linear gradients are thought to have a better resolution compared to step gradients. However, their reproducible production is tedious and requires special equipment. Therefore, we established a method to generate a step gradient with relatively small differences between the individual steps (see step 3.5). We speculated that the step gradient would be balanced upon centrifugation, resulting in an almost linear gradient. Strikingly, the determination of the sucrose concentrations of the individual fractions after centrifugation using a refractometer revealed that the step gradient indeed turned virtually linear after 12 h of centrifugation at 200,000 x g (Figure 3).
The importance of mild sonication is evident in these representative results. In mild sonication conditions (Figure 4A), the mitochondrial outer membrane marker Tom40 was enriched in the early low-density fractions (Fraction No. 4) and was virtually absent in the later ones. The mitochondrial inner membrane marker Tim17 was concentrated in high-density fractions (Fraction No. 17). In contrast, the contact site protein Mic60 accumulated in fractions of intermediate sucrose concentration (Fraction No. 12-14), indicating the successful generation and subsequent separation of the various membrane vesicles. In contrast, with harsher sonication conditions, the mitochondrial outer membrane marker Tom40 could be detected in lower fractions of the gradient (Fraction No. 14 and Fraction No. 17, Figure 4B). Additionally, there was a significant amount of Tim17 in fractions of intermediate density (Fraction No. 13 and Fraction No. 14, Figure 4B). The unspecific accumulation of outer and inner membrane markers in fractions of intermediate density indicates the artificial formation of mixed membranes, which inhibits the identification of candidate proteins.
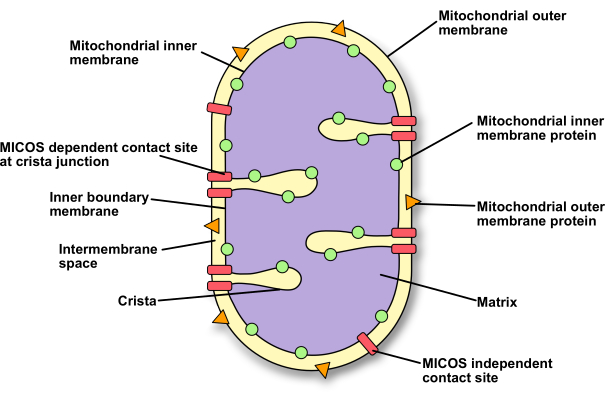
Figure 1: Schematic representation of the mitochondrial ultrastructure. A schematic representation of a mitochondrion to show the different compartments, membranes, and protein locations. Please click here to view a larger version of this figure.
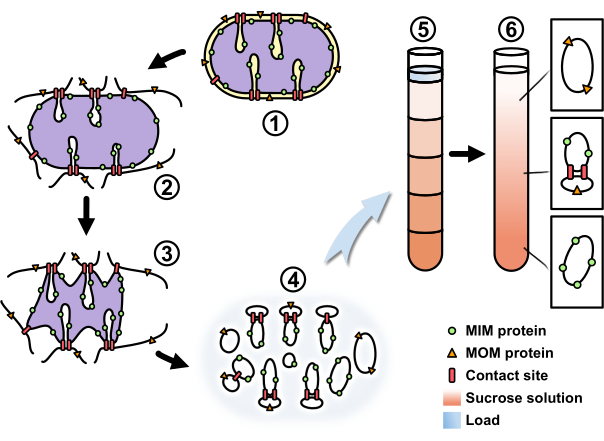
Figure 2: Workflow of mitochondrial fractionation to isolate contact site proteins. A schematic overview of the process described in the protocol. The (1) freshly isolated mitochondria were (2) osmotically swollen (3) and then shrunken. (4) The shrunken mitochondria were objected to mild sonication to generate submitochondrial vesicles. (5) The vesicle mixture was concentrated and loaded onto a sucrose step gradient. (6) Through centrifugation the mitochondrial outer membrane vesicles were enriched at a low sucrose concentration, the mitochondrial inner membrane vesicles at a high sucrose concentration, and the contact site-containing vesicles at an intermediate sucrose concentration. Abbreviations: MIM, mitochondrial inner membrane; MOM, mitochondrial outer membrane. Please click here to view a larger version of this figure.
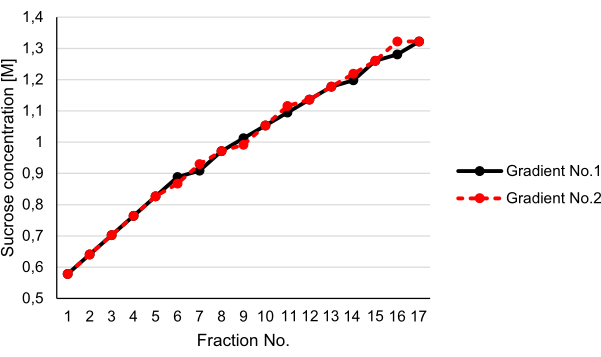
Figure 3: Generation of a linear gradient by centrifugation. Two step gradients (1.3 M, 1.13 M, then 1.02 M, 0.96 M, and 0.8 M in 20 mM MOPS and 0.5 mM EDTA, pH 7.4) were prepared in parallel, and 1 mL of 0.6 M sucrose in 20 mM MOPS and 0.5 mM EDTA, pH 7.4, was loaded onto both gradients. The gradients were subjected to centrifugation at 200,000 x g and 4°C for 12 h and harvested in 17 fractions each. The reproducibility of the method was tested by determination of the sucrose concentration of the single fractions of the individual gradients using a refractometer. Please click here to view a larger version of this figure.
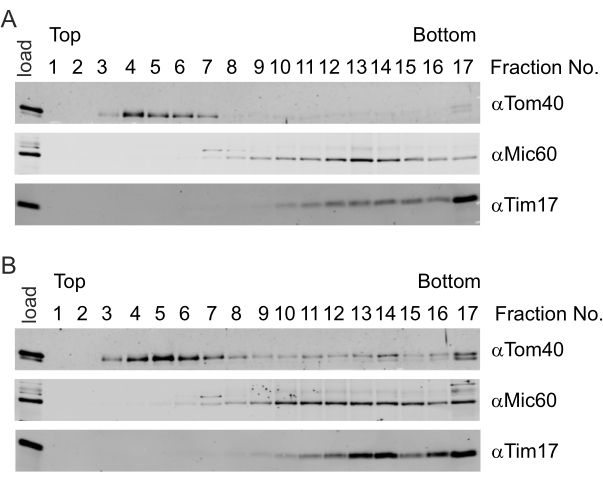
Figure 4: Isolation of contact site proteins with a well-defined ultrasonic pulse. (A,B) Freshly isolated yeast mitochondria were subjected to osmotic treatment and exposed to sonication. The generated vesicles were separated using sucrose buoyant density gradient centrifugation. The gradient was fractionated, and the proteins in the various fractions were subjected to TCA precipitation. The distribution of the marker proteins for the mitochondrial outer membrane (Tom40), the mitochondrial inner membrane (Tim17), and contact sites (Mic60) were analyzed by immunoblotting using the indicated antibodies. (A) Mild sonication (4 x 30 s, 10% amplitude) resulted in a clear separation of the outer membrane proteins (low-density fractions) and inner membrane proteins (high-density fractions). Additionally, the contact site protein Mic60 was successfully enriched in fractions of intermediate density. (B) Harsher sonication (4 x 30 s, 20% amplitude) resulted in the generation of hybrid vesicles and, thus, did not allow for a clear separation of contact sites. Please click here to view a larger version of this figure.
