


Summary
Dopamin udskiftning farmakoterapi anvendelse af L-DOPA er den mest almindeligt anvendte symptomatisk behandling af Parkinsons sygdom, men ledsaget af bivirkninger, herunder ufrivillige unormale bevægelser betegnet dyskinesi
Abstract
MALDI billeddannelse massespektrometri (IMS) er en kraftig metode til at fremme den rumlige analyse af molekylarter i biologiske vævsprøver 2 (fig. 1). En 12 um tyndt vævssnit er dækket med en MALDI-matrix, hvilket letter desorption og ionisering af intakte peptider og proteiner, som kan påvises med en masseanalysator, typisk under anvendelse af en MALDI TOF / TOF-massespektrometer. Generelt flere hundrede toppe kan vurderes i en enkelt rottehjerne vævssnittet. I modsætning til almindeligt anvendte billeddiagnostiske teknikker, ikke denne fremgangsmåde ikke kræver forudgående kendskab til molekyler af interesse og giver mulighed for uovervåget og omfattende analyse af flere molekylære arter og samtidig opretholde høj molekylær specificitet og følsomhed 2. Her beskriver vi en MALDI IMS fremgangsmåde til at belyse region-specifikke fordeling profiler neuropeptider i rottehjernen i en dyremodel for Parkinsons sygdom (PD).
PD er en almindelig neurodegenerativ sygdom med en prævalens på 1% for personer over 65 år 3,4. Den mest almindelige symptomatisk behandling er baseret på dopamin udskiftning under anvendelse af L-DOPA 5. Men dette er ledsaget af alvorlige bivirkninger, herunder ufrivillige abnorme bevægelser, kaldet L-DOPA-inducerede dyskinesier (LID) 1,3,6. En af de mest fremtrædende molekylære ændringer i låget en opregulering af opioidet precursor prodynorphin mRNA 7. De Dynorphin peptider modulere neurotransmission i hjernen områder, der hovedsageligt involveret i bevægelse kontrol 7,8. Imidlertid har til dato de nøjagtige opioide peptider, der stammer fra forarbejdning af neuropeptid forløber ikke blevet karakteriseret. Derfor har vi anvendt MALDI IMS i en dyremodel af eksperimentel Parkinsons sygdom og L-DOPA-induceret dyskinesi.
MALDI billeddannelse massespektrometri viste sig at være særlig fordelagtig med hensyn til neuropeptid characterization, idet almindeligt anvendte antistof-baserede fremgangsmåder er rettet mod kendte peptidsekvenser og tidligere observeret post-translationelle modifikationer. Derimod MALDI IMS kan opklare nye peptidbaserede forarbejdningen af produkterne, og dermed afsløre nye molekylære mekanismer i neuropeptid modulering af neuronal transmission. Medens den absolutte mængde af neuropeptider ikke kan bestemmes ved MALDI IMS, den relative forekomst af peptid ioner kan afgrænses af massespektre, hvilket giver indsigt skiftende niveau inden for sundhed og sygdom. I eksemplerne præsenteret her, blev topintensiteterne af dynorphin B, a-neoendorphin og substans P fundet at være signifikant forøget i dorsolaterale, men ikke dorsomedial, striatum af dyr med alvorlig dyskinesi involverer ansigtet, krop og orolingual muskler (fig. 5). Desuden afslørede MALDI IMS en sammenhæng mellem dyskinesi sværhedsgrad og niveauer af des-tyrosin alpha-neoendorphin, der repræsenterer en hidtil ukendt mekanisme funktionel inactirelse af dynorphiner i striatum, som fjernelsen af N-terminale tyrosin reducerer dynorphin s opioid-receptor-bindingskapacitet 9. Dette er den første undersøgelse om neuropeptid karakterisering i låget ved hjælp af MALDI IMS og resultaterne fremhæve potentiale teknik til anvendelse i alle områder af biomedicinsk forskning.
Protocol
Protokollen justeres med henblik på statistisk analyse af MALDI IMS data fra flere rottehjerne sektioner, typisk 20-30 sektioner, og består af fem forskellige trin, der omfatter vævspræparat, matrix anvendelse MALDI-TOF MS-analyse, data evaluering og neuropeptid identifikation. Procedurerne beskrevet og beskrevet mere detaljeret nedenfor:
1. Vævspræparat
Denne procedure omfatter indsamling af de respektive vævsprøver samt væv sektionering for IMS analyse. Et særligt formål med protein og peptid analyse er at undgå proteolytisk nedbrydning. Derfor er det nødvendigt at arbejde hurtigt og flittige i løbet væv dissektion.
- Sacrifice rotter (typisk 250-300 g) ved halshugning, fjerne rottehjernen inden for en maksimal post-mortem tid <30'erne og fastfrysning af pulveriseret tøris, før overførsel til -80 ° C fryser. Hurtigere frysning ved anvendelse af flydende nitrogenøge risikoen for microtears i hjernevæv, som vil påvirke matrix krystallisation og derved reducere MS kvalitet (fig. 2D). Hele hjerner kan opbevares i flere år før afskæring uden tab af MS signalkvalitet.
- Skåret frosne væv på en kryostat mikrotom til 12 um skiver og tø-monterede vævssnit på ledende MALDI objektglas (indiumtinoxid coatede objektglas, Bruker Daltonics) eller MALDI target (fig. 2A-C).
- Tørre dele for 15 min under vakuum og opbevares objektglas ved -80 ° C indtil yderligere anvendelse. Vævssnit bør analyseres inden for den kortest mulige tid efter sektionering, selv hvis den opbevares ved -80 ° C. Vi finder, at MS signal kvalitet vil blive mærkbart reduceret efter et års opbevaring. For at reducere oxidation af proteiner og peptider, kan luften i lagerbeholderen erstattes med en inert gas (f.eks argon eller nitrogen).
2. Matrix ansøgning
Matrixen anvendelsetrin har en væsentlig indvirkning på spektrum kvalitet og kræver optimering af flere parametre afhængigt af arten af vævet såvel som analytten af interesse. Disse faktorer indbefatter kemiske parametre, såsom den type matrix, matrix koncentration, pH, væv vask og organiske modifikatorer samt instrumentale indstillinger, herunder deponeringen volumen lateral opløsning og antallet af aflejringer 10 (fig. 2D). Til store eksperimenter, er det af stor vigtighed at reducere variansen, for eksempel ved anvendelse af matrixen for alle sektioner i en dag og ved den samme operatør. Selvom der er mange strategier til anvendelse matrix opløsning, såsom ved sublimering eller ved hjælp af spray, automatiseret aflejring af arrays af små matrix dråber, 100-150 picoliter i størrelse, er blevet anvendt med succes til analyse af små proteiner og neuropeptider i forskellige væv , herunder hjerne § 9, 10,11, 12, 13.
- Optø sektionerne i en ekssikkator i 1 timevores.
- Sørg for, at forsøget er blindet af en anden person end brugeren. Re-mærke alle prøver.
- Vask sektioner 1x i 70% ethanol (EtOH, ved stuetemperatur, RT) i 10 sekunder og to gange i 95% EtOH (RT) i 10 sek. For store eksperimenter udføres vasken for alle glas glider sammen ved hjælp af en kuvette for at minimere variation.
- Tørre sektioner i en ekssikkator i 10 minutter.
- Vurdere vævssnit under et mikroskop, og kontrollere, om væv forvrængning, microtears og små revner, der vil forringe MALDI MS kvalitet (Fig. 2D).
- Fremstil frisk matrix opløsning bestående af 50 mg / ml DHB i 50% methanol, 10% 150 mM ammoniumacetat (Amac) og 0,3% trifluoreddikesyre (TFA) i vand.
- Matrix ansøgning udføres ved diskrete dråbe udfældning i et rektangulært mønster med en kemisk inkjet printer (CHIP, Shimadzu). Det første skridt er at optimere de eksperimentelle parametre matrix ansøgning om neuropeptid analyse including antallet af dråber per passage, antallet af passager. Dette eksperiment udføres ved at anvende flere matrix-arrays med forskellige anvendelsesområder parametre på samme vævssnittet, samtidig med at sikre, at hver række dækker lignende områder af hjernen, som hjernebjælken, cortex og striatum. Det samme eksperiment skal udføres, hver gang parametre ændres, herunder forskellige hjernens struktur forskellige matrixer rettet mod specifikke analytter, og hvis forskellige matrixmaterialer opløsningsmidler er behov for ekstraktion af specifikke analytter.
- Scan glasset diasholderen med vævssnittet og tilpasse holderen. Definer dit array for matrix program på vævssnittet og angiv rumlig opløsning, dvs stedet at spotte afstand. Anvend matrix ved hjælp af optimerede protokol om den kemiske inkjet printer. Til dette eksperiment har vi brugt en optimeret protokol til peptid billeddannelse med følgende udskrivning parametre: 10 dråber (100 ul / drop), 10 ansøgning passerer, og et sted at spotte distance på 300 um.
- Scan endelige matrix spottet sektioner og gem billedet for registrering før MALDI dataopsamling (trin 3,4).
- Opbevare delene indtil yderligere anvendelse i en ekssikkator under vakuum.
3. MALDI MS dataindsamling og behandling
MS-analyse af neuropeptider udføres på et MALDI flyvetid instrument (Ultraflex II, Bruker Daltonics, Tyskland), der reflektor tilstand ved hjælp af software assisteret datafangst fra hver enkelt matrix plet 14. Derfor præcis rumlig undervisning er af afgørende betydning. Det er vigtigt, at MALDI optimering, indkøb og navnlig målet om registrering forsøgene udføres af den samme operatør, der skal helst være blindet for de eksperimentelle grupper. I stor skala eksperiment med flere glasplader, kan MALDI eksperimenter udføres af en operatør, mens en anden person, der betjener kemiske inkjet printer.
- Indlæse objektglas i massespektrometret.
- Se kalibrering af MALDI overtagelsesmetoden med en lav molekylvægt standardkalibreringskurve blanding (Bruker Daltonics).
- Optimer erhvervelse parametre.
- For at optimere MS signal og til at undgå ablatere matricen fra tilstødende matrix aflejringer, bør antallet af laseren og den optimale vægt på væv bestemmes.
- Laserenergien er indstillet til at sikre maksimal MS kvalitet fra så mange matrix aflejringer som muligt uden at hæve basislinien, reducere top opløsning eller mætte detektoren.
- Vurdere det maksimale antal billeder pr matrix stedet indtil kun støj opdages, ofte 1000-2000 skud. Beregn antallet af billeder der skal akkumulerede, og antallet af skud erhvervet før laser position inden for en plet skulle ændre sig. For at udtage prøver hver matrix plet jævnt vi akkumulerer 600 skud i 25 skud trin, til et samlet antal på 24 trin med et random bevægelsesmønster, fra hver matrix aflejring.
- Registrer scanning af alle plettede sektioner til motoriske koordinater for MALDI scenen ved hjælp af FlexImaging (v.2.0), 10 og udføre dataopsamling i batch-mode med AutoXexuteBatchRunner.exe software.
- Behandl hver enkelt spektre ved hjælp af baseline subtraktion (konvekse V3), udjævning og ekstern kalibrering (valgfrit), efterfulgt af eksport som en ascii-fil (*. Dat, *. Txt eller *. Csv-format). 15
4. Datavurdering
Endelig datavurdering omfatter data efterbehandling og datareduktion ved at fokusere udelukkende på peak oplysninger, efterfulgt af statistisk analyse.
- Som et indledende skridt blev MALDI IMS sektioner evalueret for overnormalization effekter. Dette kan let opnås ved anvendelse af data visualiseringsværktøjer såsom FlexImaging (Bruker Daltonics) eller BioMap (Novartis). Som et indledende skridt de samlede ion billeder evaluated før total ionstrøm (TIC) normalisering, fulgt af en manuel undersøgelse af enkelt ion distribution billeder af forskellige fremtrædende peptidtoppe. Kig efter karakteristiske peak intensitet distributioner, og hvis de er relateret til væv funktioner (erstatning), pletblødning kvalitet eller normalisering effekter (Fig. 3).
- Afgrænse områder af interesse (f.eks striatum) i henhold til histologiske træk og eksportere de tilsvarende spektre i ASCII filformat. Fortrinsvis kan normalisering af spektre til den samlede ionstrøm (TIC) udføres på dette stadium.
- Import ascii-filer i en database håndtering software såsom Oprindelse (v.8.1, Originlab), MATLAB (MathWorks, Natick, MA, USA) eller R 16. Peak detektering kan udføres ved hjælp af peak finde værktøjer, der indgår i softwaren, for eksempel "peak analyse" oprindelse eller "mspeaks" i Matlab. Eksporter peaklists fra alle spektre som én tabulatorsepareret tekstfil.
- For at bestemme bin grænser for opdaget peptidtoppe, er Binning analyse udført ved hjælp af passende software-værktøjer (f.eks pbin 17) eller in-house skrevet manuskripter til MATLAB eller R. Her den enkelt tekstfil, der indeholder alle peak hentet data indlæses i softwaren og parametrene for maksimal grænse bestemmelse er specificeret såsom hvor ofte en top bør være til stede i spektre for at være relevant for eksperimentet. For eksempel indeholder eksperimentet 2 grupper af dyr, 5 dyr i hver gruppe, og 100 spektre er opsamlet fra hvert dyr og region af interesse. Antage en spids er potentielt interessant, hvis den er til stede i i det mindste størstedelen af dyrene i en gruppe (3/5) og mindst halvdelen af spektrene af disse dyr (3x50 = 150 spektre), vil dette give en samlet procentdel 15% for 150 positive spektre ud af den samlede 1000 (2x5x100) spektre. Ved hjælp af pbin værktøjet dette trin giver en enkelt binrange fil der indeholder alle bin bredder bestemt ud fra de opsamlede data. For at kontrollere, at bin grænserer passende, er det nemt at visualisere de spande i Origin sammen med de oprindelige spektre spor.
- Peak-området integration kan reducere variansen, der er vigtigt for statistisk analyse. Vi bruger en in-house skrevet manuskriptet til R for at beregne arealet under kurven mellem peak-grænser bestemt i trin 4. Integrerede toparealer importeres til MS Excel (v.2007) og statistisk analyse ved hjælp af ikke-parametrisk uparret test ved brug af SAM værktøjet udføres 18.
5. Peptid identifikation
Sekvens verifikation af de observerede peptid identitet er afgørende for at indgå biologiske relevans. Den mest nøjagtige metode omfatter sand top-down-bestemmelse direkte fra vævet med peptid fragmentering ved hjælp af tandem massespektrometri (MS / MS), skønt høje peptidkoncentrationer kræves til denne type analyse 12,13. For lav rigelige peptider eller flere peptider med closEM / z-værdier (± 0,5%), er on-vævsanalyse svækket og fra væv analyse under anvendelse af en peptidomic strategi anvendes som omfatter ekstraktion, adskillelse og MS baseret identifikation af endogene neuropeptider. For eksperimentet præsenteret her, var fokuseres på opioid-peptid detektion, hvilket udgør en særlig udfordring, da disse peptider er meget lav overflod sammenlignet med andre neuropeptider i spektrene. Desuden er disse peptider er ikke polære hvilket gør dem relativt hydrofile og vanskelige at opbevare med fælles peptid ekstraktion og separationsteknikker .. Derfor har vi anvendt en tidligere rapporteret protokol for væv udvinding og opioid peptid præfraktionering i kombination med standard LC-MS/MS baseret peptid identifikation 9,19.
- Indsamle koronale sektioner af målstrukturerne af interesse nucleus accumbens, NAc; putamen caudalis, CPU). Montere frosne rottehjerne i en kryostat mikrotom og fjerne omgivende hjerne materiale (cortex, septum, corpus callosum) med en skalpel. Saml sektioner (30 um, n = 50) af dissekeret NAC og CPU og tø montere NAC og CPU dele af afsnittene om forskellige objektglas.
- Ekstrahere peptider off væv ved tilsætning af 100 pi 5% ACN/0.1% TFA, inkuberes i to minutter og opsamles i Eppendorf lav proteinbinding rør. Gentag dette trin to gange.
- Udføre peptid præfraktionering ved hjælp af en stærk kationbytter-chromatografi under anvendelse trinvis (n = 4) eluering ved forøget ionstyrke 19. Tørre ned prøver under vakuum under anvendelse af en SpeedVac koncentrator.
- Analysere peptid fraktioner ved hjælp af nanoflow C18 omvendt fase væskekromatografi (1100, Agilent Technologies, Santa Clara, CA) interfaced til elektrospray tandem-massespektrometri (LC-MS/MS). MS blev udført på en hybrid lineær iontrap / Fouriertransformation ion-cyklotron-resonans (FTICR) instrument (LTQ FT 7T, Thermo Scientific, Waltham, MA,) Peptid fulscan spektre (m / z 150-2000) Blev erhvervet med FTICR analysatoren ved høj masse opløsning (100000) efterfulgt af efterfølgende opsplitning af de 5 mest intense peptidtoppe i iontrap ved hjælp af kollision induceret dissociation (CID) 9.
- Peptidsekvens identifikation udføres ved database matchning og kan suppleres med de-novo-sekventering analyse. For databasesøgning, der er kommercielt tilgængelige søgemaskiner (Mascot, XTandem eller Protein Prospector) ansat 20. De søger typisk udføres i forhold til databaser, der indeholder sekvenser af kendte eller forudsagte neuropeptider og sekvenser af neuropeptid præcursorproteiner 21.
6. Repræsentative resultater
MALDI billeddannelse massespektrometri af striatale vævssnit som fremstillet ifølge protokollen beskrevet her resulterede i detektion af mere end 1000 toppe svarende til cirka 300 monoisotopic molekylarter (gennemsnit spektre vistei fig. 1). Datavisualisering for fremtrædende molekylære ion-toppe blev opnået ved hjælp af Flex Imaging software og viste karakteristiske maksimal intensitet distributioner, der er godt i tråd med anatomiske træk (Fig.3). Et yderligere træk ved MALDI IMS er dens relative god reproducerbarhed. I dette eksperiment var den samlede varianskoefficient for topintensiteterne af alle detekterede molekylarter 30%, men mange toppe viste meget lave variationer og høj reproducerbarhed inden behandlingsgrupper (fig. 4). De relative maksimale intensitet data fra fire forskellige regioner af interesse, herunder dorsolaterale og dorsomedial del af både læderet og intakt striata blev udsat for statistisk analyse. For at korrigere for multiple sammenligninger samtidigt, blev statistiske analyser udført ved hjælp af ikke-parametrisk test under anvendelse af SAM værktøjet 18. De mest fremtrædende ændringer fundet i dorsolaterale del af dopamin-denerverede, Parkinson striatum. Her si gnificant ændringer i de forskellige behandlingsgrupper blev observeret for to dynorphin peptider; dynorphin B og alpha-neoendorphin (Fig.5). I detaljer blev en relativ forøgelse af både dynorphin topintensiteter med 50-60% observeret i høje dyskinetiske dyr sammenlignet med lave dyskinetiske dyr og læsioner kontroller (p <0,05, F (2, 15) = 12,8 DynB, F = 5,7 aNeo; Figur 5).

. Figur 1 Gennemsnitlig MS sporer fra to nært beslægtede regioner i striatum, putamen caudalis (CPU) og nucleus accumbens (NAc). De to regioner viser forskellige MS profiler med nogle molekylære arter unikt udtryk i en region, eller på forskellige peak intensitet niveauer (indsæt, m / z 2028). Den rumlige fordeling mønster af hver top kan visualiseres ved hjælp af specialiserede billedbehandlingssoftware (nederste panel).
2.jpg "/>
Figur 2 (a) Hjernen er monteret på en kryostat spændepatron med et omsluttende medium (OTC; pil). Drages der omsorg for, at OTC ikke forurener det område af hjernen er snit, eftersom OTC årsag ion suppression af peptider. (B, C) Tynde snit (≈ 12 um tykkelse) er tø-monteret på MALDI kompatible objektglas og tørret i nogle få sekunder for at undgå frostskader som det ses i C. (D) Microtears kan være vanskeligt at detektere det blotte øje , men forringer MALDI-matrix krystallisation og udslette MALDI MS signal. Den samme sektion farves med cresylviolet viser microtears og revner (nederst til højre mikrofotografi).

Figur 3. Det første skridt i data evaluering er at visualisere flere forskellige toppe på tværs af masseinterval analyseret (AI). Her blev striatale sektioner fra 9 mus afbildet med MALDI MS. Visualisering af den samlede gennemsnitlige inden aktuelle vil afsløre områder iøjnefaldende høje eller lave ion intensiteter (pile). Disse områder kan blive påvirket af over-eller-normaliseringsreferencerne effekter og fordreje dataanalyse gå på kompromis med resultaterne. Dårlig anatomiske definition af peak-distributioner afslører sektioner med generelt lav peak signal-til-støj, for eksempel § § 3 og 9, toppe F gennem I.

Figur 4. MS reproducerbarheden mellem behandlingsgrupper kan vurderes ved at beregne den gennemsnitlige MS spor og standardfejlen for hver m / z-værdi (indsætter, m / z 722 og 1749). God reproducerbarhed sikrer validt statistisk analyse.
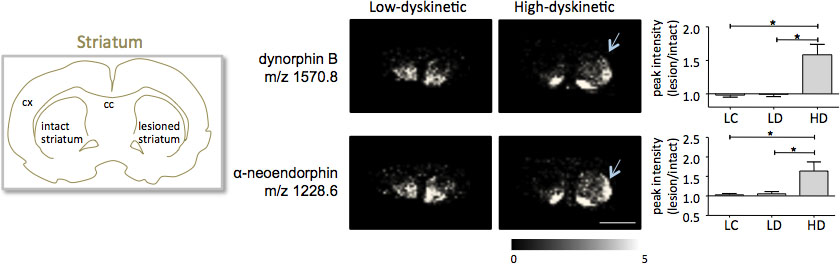
Figur 5. Dynorphin B og alfa-neoendorphin Topintensiteterne er signifikant forøget i 6-OHDA-Læderede, Parkinson, striatum af høj-dyskinetiske dyr (HD, pile) i forhold til lav-dyskinetiske (LD) og læsion kontrolgruppen (LC). Peptid topintensiteter udtrykt som gennemsnit fold-ændringen af intakte side ± SEM (læsion / intakt side). * P <0,05; cx cortex; cc corpus callosum. Målestokken 5 mm.
Discussion
Der er adskillige fordele ved anvendelse af MALDI billeddannelse massespektrometri i studiet af neuropeptider. En neutral analyse af MS-data, kan afsløre, at kun specifikke hjernekerner, eller som i de resultater, som præsenteres her, hvor kun dorsolaterale del af striatum er forbundet med en vis patofysiologisk tilstand. Ved at fastholde den geografiske oplysninger er det så muligt at redefinere områder af interesse at udføre statistisk analyse med højere følsomhed og lavere variabilitet sammenlignet med en analyse af hele hjernen sektioner eller ved hjælp af traditionelle peptidomics undersøgelser om peptid ekstrakter. Desuden er det vigtigt at indse MALDI IMS let kan detektere hidtil ukendte post-translationelle modifikationer, men strukturelle analyser skal følge for at bestemme de nøjagtige aminosyrepositioner, der er modificeret.
Almindelige faldgruber i at visualisere en MALDI IMS-data omfatter kortlægning til det maksimale intensitet til en lineær optisk skala fra blACK (0%) til farve (100%) for hver sektion i den eksperimentelle serie (fig. 3) i stedet for kortlægning af alle sektioner af en fælles absolut skala, hvor 100% er det maksimale intensitet af alle sektioner (fig. 5) . Sidstnævnte metode giver mulighed for sammenligning af gruppe data og visualisering af forskelle mellem behandlingsgrupper.
En væsentlig hindring i MALDI IMS analyse er tildeling af peptider til specifik masse toppe. På væv tandem-massespektrometri er nogle gange muligt, men ofte viser sig ganske vanskeligt 13,14. Vi finder, at en mere traditionel fremgangsmåde, herunder en præparativ fraktionering på en stærk kationbytter-chromatografi, efterfulgt af omvendt fase LC-MS/MS kan anvendes med succes sekvens mange neuropeptider og især opioide peptider. Det er stadig ikke ualmindeligt at opnå god kvalitet MS / MS-spektre, som ikke matcher nogen database poster ved hjælp af almindelige søgemaskiner såsom MASCOT. I disse tilfælde de novo-sekventering ved håndkraft, er ONLy mulighed. Det endelige bevis for top identitet kan opnås ved MALDI IMS af vævssnit fra en passende knockout-mus, men dette er ikke altid til rådighed eller er muligt. Et alternativ er at validere de resultater, ved en diametralt anden metode, for eksempel ved western immunoblotting eller immunhistokemi. Dette kan ofte indbefatte fremkaldelse af antistoffer, og en betydelig mængde arbejde validere de nye antistoffer.
Den generelle strategi, der præsenteres i denne protokol er optimeret til store neuropeptid MALDI IMS eksperimenter, herunder flere sektioner og eksperimentelle betingelser. Protokollen er optimeret specielt til opioide peptider og vil få stor betydning i fremtidige undersøgelser, som ansat i forskellige forskningsområder, herunder mekanismerne bag smerten og den endogene reaktion på lægemidler i afhængighed.
Disclosures
Forfatterne har ikke noget at afsløre.
Acknowledgments
Vi takker Hanna Warner for at bidrage data til figur 3 og professor Jonas Bergquist for værdifulde input. Det svenske Forskningsråd (Grant 522-2006-6416 (MA), 521-2007-5407 (MA), Den Åke Wiberg Fond (MA, JH), Det Kongelige svenske videnskabsakademi (MA, JH), og det svenske Kemisk Society (JH) er modtaget med tak for økonomisk støtte.
References
- Obeso, J. A., Olanow, C. W., Nutt, J. G. Levodopa motor complications in Parkinson's disease. Trends Neurosci. 23, S2-S7 (2000).
- Caprioli, R. M., Farmer, T. B., Gile, J. Molecular imaging of biological samples: localization of peptides and proteins using MALDI-TOF MS. MALDI-TOF MS. Anal. Chem. 69, 4751-4760 (1997).
- Obeso, J. A. The evolution and origin of motor complications in Parkinson's disease. Neurology. 55, S13-S20 (2000).
- O, W. H. Noncommunicable Diseases and Mental Health Cluster, Noncommunicable Disease Prevention and Health Promotion Department, Ageing and Life Course. Active Ageing: A Policy framework. , (2002).
- Schapira, A. H. Movement disorders: advances in cause and treatment. Lancet Neurology. , 6-7 (2010).
- Obeso, J. A., Rodriguez-Oroz, M. C., Rodriguez, M., DeLong, M. R., Olanow, C. W. Pathophysiology of levodopa-induced dyskinesias in Parkinson's disease: problems with the current model. Ann. Neurol. 47, S22-S32 (2000).
- Cenci, M. A., Lee, C. S., Bjorklund, A. L-DOPA-induced dyskinesia in the rat is associated with striatal overexpression of prodynorphin- and glutamic acid decarboxylase mRNA. Eur. J. Neurosci. 10, 2694-2706 (1998).
- Andersson, M., Hilbertson, A., Cenci, M. A. Striatal fosB expression is causally linked with l-DOPA-induced abnormal involuntary movements and the associated upregulation of striatal prodynorphin mRNA in a rat model of Parkinson's disease. Neurobiol Dis. 6, 461-474 (1999).
- Hanrieder, J. Alterations of striatal neuropeptides revealed by imaging mass spectrometry. Molecular & Cellular Proteomics. , (2011).
- Cornett, D. S., Reyzer, M. L., Chaurand, P., Caprioli, R. M. MALDI imaging mass spectrometry: molecular snapshots of biochemical systems. Nat. Methods. 4, 828-833 (2007).
- Ljungdahl, Imaging Mass Spectrometry Reveals Elevated Nigral Levels of Dynorphin Neuropeptides in L-DOPA-Induced Dyskinesia in Rat Model of Parkinson's Disease. PLoS ONE. 6, e25653 (2011).
- Groseclose, M. R., Andersson, M., Hardesty, W. M., Caprioli, R. M. Identification of proteins directly from tissue: in situ tryptic digestions coupled with imaging mass spectrometry. J. Mass. Spectrom. 42, 254-262 (2007).
- Andersson, M., Groseclose, M. R., Deutch, A. Y., Caprioli, R. M. Imaging mass spectrometry of proteins and peptides: 3D volume reconstruction. Nat. Methods. 5, 101-108 (2008).
- Deininger, S. -O. Imaging Mass Spectrometry. Setou, M. , Springer. Japan. 199-208 (2010).
- Norris, J. L. Processing MALDI Mass Spectra to Improve Mass Spectral Direct Tissue Analysis. Int. J. Mass. Spectrom. 260, 212-221 (2007).
- Ihaka, R., Gentleman, R. R. A Language for Data Analysis and Graphics. Journal of Computational and Graphical Statistics. 5, 299-314 (1996).
- Mass Spectrometry Binning Software GAB. , Vanderbilt Center for Quantitative Sciences. Nashville, TN. Available from: http://www.vicc.org/biostatistics/software.php (2012).
- Tusher, V. G., Tibshirani, R., Chu, G. Significance analysis of microarrays applied to the ionizing radiation response. Proc. Natl. Acad. Sci. U.S.A. 98, 5116-5121 (2001).
- Bergstrom, L., Christensson, I., Folkesson, R., Stenstrom, B., Terenius, L. An ion exchange chromatography and radioimmunoassay procedure for measuring opioid peptides and substance P. Life. Sci. 33, 1613-1619 (1983).
- Falth, M. Neuropeptidomics strategies for specific and sensitive identification of endogenous peptides. Mol. Cell. Proteomics. 6, 1188-1197 (2007).
- Falth, M. SwePep, a database designed for endogenous peptides and mass spectrometry. Mol. Cell. Proteomics. 5, 998-1005 (2006).
