


Summary
En rask
Abstract
Screening forbindelser for in vivo aktivitet kan brukes som et første skritt for å identifisere kandidater som kan utvikles til farmakologisk agenter 1,2. Vi utviklet en roman nanoinjection / elektrofysiologi analysen som tillater påvisning av bioaktive regulerende effekt av forbindelser på funksjonen til en neuronal krets som medierer flukten respons i Drosophila melanogaster 3,4. Vår in vivo-forsøk, som bruker Drosophila Giant Fiber System (GFS, figur 1) tillater screening av ulike typer forbindelser, for eksempel små molekyler eller peptider, og krever bare minimale mengder å lokke fram en effekt. I tillegg tilbyr Drosophila GFS et stort utvalg av potensielle molekylære mål på nerveceller og muskler. The Giant fiber (GFS) synapse elektrisk (Gap veikryss) samt kjemisk (kolinerge) på en ytre Synapsing Interneuron (PSI) og Tergo Trochanteral Muscle nervecellen (TTMn) 5 6. Til slutt, den nevromuskulære knutepunktene (NMJ) av TTMn og DLMn med hopp (TTM) og fly muskler (DLM) er glutamatergic 7-12. Her viser vi hvordan du injiserer nanoliter mengder av et stoff, mens skaffe elektrofysiologiske intracellulære opptak fra Giant Fiber System 13, og hvordan å overvåke virkningene av det sammensatte på funksjonen til denne kretsen. Vi viser spesifisitet av analysen med methyllycaconitine citrat (MLA), en nAChR antagonist, som forstyrrer PSI til DLMn tilkobling, men ikke GF å TTMn tilkobling eller funksjonen av NMJ på hopp eller fly muskler.
Før du begynner denne videoen er det viktig at du nøye se og bli kjent med Jove video med tittelen "Elektrofysiologiske Opptak fra Giant Fiber Pathway av D. melanogaster "fra Augustin et al 7, som video som presenteres her er ment som en utvidelse til denne eksisterende teknikken. Her bruker vi elektrofysiologisk innspillinger metode og fokus på detaljer bare om tilsetting av de sammenkoblede nanoinjections og overvåking teknikk.
Protocol
1. Elektrofysiologi Rig Set-up
- Den nødvendige utstyret for elektrofysiologi riggen Oppsettet er beskrevet i detalj av Augustin et al. i denne Journal 14. Vennligst referer til denne artikkelen for en detaljert forklaring av elektrofysiologiske apparater som trengs.
- Endre tidligere beskrevet elektrofysiologi rigg set-up 14 ved å legge en sjettedel micromanipulator, som holder nanoinjector. For enkel tilgang til hodet på dyret det skal plasseres mellom de to stimulerende elektrode micromanipulators som vist i Figur 2.
- Før du starter forsøket, sørge for at du har en komfortabel rekkevidde på alle aksene i rotasjon med alle micromanipulators, og at alle elektroder samt injeksjon mikropipette (se trinn 2) kan nå dyret.
2. Nanoinjection Set-up
- Den nanoinjection set-up krever en Nanoliter2000 (World Precisipå Instruments, Sarasota, FL, USA) eller tilsvarende type injektor som gjør kontrollerte injeksjoner i nanoliter mengder.
- Forbered injeksjonsnål hjelp av glasset mikropipetter følger med injektoren ved å trekke dem til en motstand på 80-100 MΩ med en elektrode avtrekker.
- For glatte injeksjoner det er nødvendig å bevel de mikropipetter til en 11-17 mikrometer åpning i en 45 graders vinkel (figur 3).
- Sakte etterfylle injeksjonen mikropipette med syntetisk olje med en Hamilton sprøyte som instrueres av Nanoliter2000 manuelle, slik at ingen luftbobler er til stede.
- Nøye sikre mikropipette på nanoinjector og forberede det å laste det sammensatte ved å tømme overflødig olje som instrueres av Nanoliter2000 håndboken.
- Plasser injektoren på micromanipulator og laste det sammensatte som instruert i Nanoliter2000 manualen. Pass på at spissen på mikropipette ikke bryter i løpet av denneprosedyre.
- Still inn ønsket mengde nanoliters som skal injiseres i injektoren kontrollboks som instrueres av Nanoliter2000 håndboken. Vær oppmerksom på at totalbeløpet injiserte ikke bør overstige 100 nl. Vi fant at større mengder saltvann kontroll løsninger kan påvirke funksjonen til GFS kretsen.
- Det er avgjørende å ha injektoren koblet fra styreboksen under innspillingen oppkjøpet med unntak av injeksjon selv, siden strømforsyningen til nanoinjector forstyrrer opptak, som er synlig som bakgrunnsstøy (figur 4). Men ikke koble strømforsyningen til kontrollboksen som det vil re-sette den.
3. Drosophila melanogaster Forberedelse
- Anesthetize 2 til 6 dager gammel fluer med CO 2 eller på is som tidligere beskrevet 14,15.
- Når immobile, bruk en pinsett til å overføre dyret til en liten tallerken with myk dental voks ved å plukke den opp fra bena. Vær oppmerksom på at en mannlig bananflue veier ca 1,0 mg og en kvinnelig bananflue veier ca 1,2 mg, og dermed sammensatte til kroppsvekten forholdet er annerledes i mannlige versus kvinnelige fluer. Derfor anbefales det å bruke bare ett kjønn for eksperimentene.
- Som tidligere beskrevet 14,15, forsiktig montere flua dorsal side opp og sikre at brystkasse og hode er immobilisert med myk dental voks plassert rundt i kroppen. Forsiktig spre vingene ut, slik at de lå vinkelrett på thorax (figur 2, C). Flua bør monteres med så lite skade som mulig.
4. Sammenkoblede Nanoinjection / elektrofysiologi
- Plasser montert flue på elektrofysiologi riggen med hodet mot stimulerende elektrodene.
- Spidde dyret med tilsvarende stimulerende, jord og opptak elektroder som tidligere beskrevet 14,15 (F2 igure, C). Med mindre annet er ønsket, plasser en innspilling elektrode inn i DLM og den andre inn i TTM muskelen. Den DLM muskel ligger i thorax mellom de fremre Dorso-Sentrale hår og midtlinjen på flua. Den TTM muskelen ligger nær vingen vedlegg, mellom de bakre og fremre Supra-Alar hår på flua 7.
- Juster injeksjon mikropipette inneholder sammensatte med midten av de tre Ocelli ligger på den mediale bakre delen av hodet, men ikke injisere ennå (figur 2, C).
- Før sammensatte injeksjon få en baseline opptak av GF til TTM og GF til DLM veier i Giant Fiber System (GFS, figur 1) via hjernestimulering. For å gjøre dette, aktivere Giant fiber (GFS) med 10 tog i 10 stimuli (40-50 mV) hver gitt for en varighet på 0,03 ms ved 100 Hz med en 1 sekund forsinkelse mellom togene 14,15 (figur 4). Villtype flyr shOuld kunne følge en-til-en på denne sats på stimulering for både DLM og TTM veier. Kast flua hvis GF til DLM og GF til TTM veier ikke følger den 100 Hz stimulering i forholdet 1:1.
- Bytt til kontinuerlig stimulering av GF med enkle pulser på 1 Hz (figur 4).
- Raskt plugge injektoren i kontrollboksen. Selv om bakgrunnsstøy vil forstyrre med 1 Hz stimulering opptak, ikke avbryte det.
- Forsiktig sett injeksjon mikropipette i hodet kapsel på flua like under cuticule, og injiserer ønsket mengde sammensatte inn hemolymph på flua samtidig opprettholde en Hz stimulering (figur 4). På grunn av den åpne sirkulasjonssystemet på flua, vil hele nervesystemet bli utsatt for det sammensatte i løpet av sekunder. Selv om en bestemt injeksjonsstedet er ikke avgjørende for å levere forbindelser inn i hemolymph, finner vi den regionen i Ocelli, som er lokalisered mediale på det mest dorsal siden av hodet kapsel, for å være et praktisk nettsted som gir en enkel injeksjon som fører til rask og jevn fordeling av det sammensatte.
- Umiddelbart fjerne injeksjon mikropipette fra injeksjonsstedet og trekk injektoren fra styreboksen mens du fortsetter 1 Hz GF stimulering opp til 1 min etter injeksjon (figur 4).
- For å avdekke mer subtile effekter av forbindelser på GFS, stresse Giant fiber (GFS) med 10 tog i 10 stimuli ved 100 Hz med en 1 sekund forsinkelse mellom togene. Fortsett å teste funksjonen av GF veier med dette paradigmet hvert 5 minutter opp til 15 minutter (figur 4). Men kortere intervaller eller lengre overvåking perioder er også mulig.
- For å teste om massen har en effekt på det nevromuskulære knutepunktene (NMJs) av GFS, og muligens begrense virkningene av det sammensatte, fortsette å aktivere motoren neurons retningtly av thorax stimulering. For dette, fjerne stimulerende elektroder fra øynene og erstatte dem på fremre sider av thorax for å stimulere de motoriske nevroner med 10 tog i 10 stimuli på 100 Hz.
Merk: De elektrofysiologiske spor vises på videoen ikke samsvarer med effekten av ren fargestoff injeksjon.
5. Representative Resultater
Effekt av en antagonist på PSI til DLM synapse av Giant Fiber System
Methyllycaconitine citrat (MLA) er en nAChR antagonist som er spesifikk for α7 nAChR subtyper. PSI til DLMn synapse i GF-DLM veien er avhengig av Dα7 nAChR undertype for riktig funksjon, mens genetisk fjerning av Dα7 nAChR subtype har ingen effekt på GF-TTM veien 5,6. For å demonstrere spesifisitet og sensitivitet av analysen vår vi injisert MLA på ulike konsentrasjoner (0, 0,02, 0,04, 0.08, 0,12 ng / mg, 46 nl injisert) inn i hodet på dyret (n = 10 per sammensatt behandling; n = 15 for saltvann behandling). Bare mannlige fluer (av villtype genotype vill 10E) ble brukt, og effekten av det sammensatte ble overvåket for totalt 15 minutter etter injeksjon.
Figur 5 viser forskjellen mellom baseline opptak innhentes før injeksjonen og de innhentet etter injeksjon i respons til MLA og saltvann kontroll løsning. Vi fant at injeksjon av MLA resulterte i manglende evne til GF-DLM vei å følge en-til-en på 100 Hz ved stimulering av GFS i hjernen mens GF-TTM veien forble upåvirket. (Figur 5, øvre og midtre spor, t-test utført mellom saltvann kontroller [0 ng / mg] og ulike konsentrasjoner av MLA på hvert tidspunkt med mindre dataene er ikke-parametrisk [normalitet og lik varians testet], ellers bruker vi en Mann-Whitney Rank Sum Test. * p <0,001). Men en en-til-en response av DLM ble observert da de motoriske nevronene ble stimulert direkte (Figur 5, bunn), viser at NMJ funksjon DLM og TTM ikke påvirkes av MLA. MLA ut til å nå sitt maksimale effekt 1 minutt etter injeksjon for 0,04, 0,08 og 0,12 ng / mg MLA injisert, som ingen ytterligere signifikante endringer ble registrert i løpet av de neste 15 minuttene av testperioden. Videre nådde det sammensatte en maksimal effekt på 0,08 ng / mg siden sterkere svarene ikke ble observert med høyere dose på 0,12 ng / mg.

Figur 1. The Giant Fiber System Diagram av Giant Fiber System (GFS). The Giant fiber (GFS, vist i rødt) synapse elektrisk (Gap veikryss) samt kjemisk (kolinerge) på en ytre Synapsing Interneuron (PSI, vist i grønt) og den Tergo Trochanteral Muscle nevron (TTMn, vist i gult) 5. PSJeg DLMn (Dorsal Longitudinal Muscle nevron, vist i blått)-tilkobling er avhengig Dα7 nAChR subtype 6. Endelig nevromuskulære krysset (NMJ) av TTMn og DLMn på hopp (TTM, vist i lilla) og fly muskler (DLM, vist i lilla) er glutamatergic.
Merk: GF til PSI forbindelse er både elektrisk og kjemisk. Men i shakB mutanter (som mangler gap veikryss), kan ingen respons tas opp fra DLM ved stimulering av GFS i hjernen, viser at den kjemiske komponenten i fravær av elektriske koblinger er ikke tilstrekkelig til å fremkalle en handling potensial i PSI 5,16-18. Fordi GF til PSI forbindelse er gap junction avhengig, viser dette tallet bare GAP krysset ved synapse for enkelhet grunner.

Figur 2.
Micromanipulators set-up.
- En modifisert set-up av en tidligere utgitt protokoll 14 brukes til å passe injeksjon micromanipulator for sammenkoblede GFS innspillinger med samtidige nanoinjections. Den monteres flua preparatet er innrettet med hodet på flua mot eksperimentator. Injeksjonen micromanipulator (# 1) er plassert i fronten av eksperimentator mellom de to manipulatorer for wolfram stimulerende elektroder (2 # og # 3). De to micromanipulators for glasset opptak elektrodene (# 4 og # 5) er plassert på venstre og høyre side, henholdsvis. Det micromanipulator for wolfram jordelektroden (nr. 6) er plassert lengst bak på høyre side (vist her) eller på høyre side.
- Et nærbilde utsikt fra toppen av ordningene i de ulike elektroder og injeksjon mikropipette.
- Et riktig montert D. melanogaster spiddet med elektroder og injeksjon mikropipette klar til injeksjon. Legg merke til at dyrets kropper montert med sin brystkasse horisontalt og vingene spredt ut. Voksen er skikkelig pakket rundt kroppen hindrer at dyret beveger seg. I tillegg jordelektroden (nr. 6, i magen), glasset opptak elektrodene (# 4 og # 5, i thorax, fremhevet av mørke konturer), og de stimulerende elektroder (# 2 og # 3, en på hvert øye ) er spiddet på plass, som tidligere beskrevet 14. Injeksjonen mikropipette (# 1) er riktig justert med midten av de tre Ocelli (innringet). Innsetting av injeksjonen mikropipette bør plasseres i dette området.

Figur 3. Avskårne injeksjon mikropipette. Et diagram av en riktig beveled mikropipette vises her. Elektroden Åpningen bør skrå i en 45 graders vinkel og har en åpning mellom 11 til 17 mikrometer. En skikkelig skrå injeksjon mikropipette er avgjørende for en jevn injeksjon med minimal damage til flua.

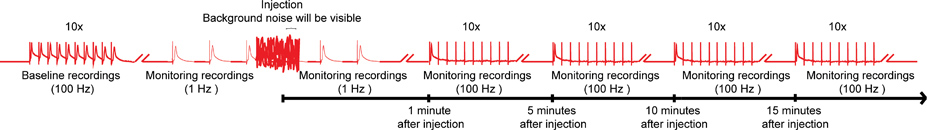
Figur 4. Generelle ordningen for nanoinjection / elektrofysiologi protokollen. En representant diagram av den generelle ordningen for nanoinjection / elektrofysiologi protokollen. Start med å skaffe en baseline opptak ved å stimulere Giant fiber (GFS) ved 100 Hz med 10 tog i 10 stimuli hver (kun ett tog vist her). Før injeksjon, begynner de 1 Hz stimuleringer ett sekund fra hverandre. Under injeksjon tid (mens injektor er koblet til kontrollboksen), vil du observere betydelig bakgrunnsstøy, men ikke avbryte innspillingene. Etter injeksjon (og injektor er koblet fra kontroll boks), fortsetter 1 Hz stimulering for ca 1 minutt. Til slutt, fortsetter å understreke GFS med 10 tog i 10 stimuli på 100 Hz og fortsette å teste funksjonen av GF veier med dette paradigmet hvert 5 minutter opp til 15 minutter. Merk: opptak were manipulert for å skape den generelle ordningen, og representerer ikke et bestemt resultat som oppnås. Ikke for å skala, blir ikke alle spor vist. Klikk her for større bilde .

Figur 5. Effektene av MLA i GFS.
- En grafisk fremstilling av virkningene av den α7 nAChR antagonist Methyllycaconitine citrat (MLA) på GF-DLM vei av bananflue på ulike konsentrasjoner (0, 0,02, 0,04, 0,08, 0,12 ng / mg n = 10 per sammensatt behandling.; n = 15 for saltvann behandling). Kun ett minutt etter injeksjon, ble en betydelig og umiddelbar virkning sett med 0,04 ng / mg MLA. En signifikant effekt ble også sett med 0.8ng/mg og 0.12ng/mg av MLA ved 100Hz stimulering av GFS. Ingen signifikant forskjell ble observert mellom saltvann kontroller og 0,02 ng / mg MLA. I tillegg er det ingen endring in effekt ble sett etter 1 minutt etter injeksjonen i den tiden testet (15 minutter). En t-test ble utført mellom saltvann kontroller (0 ng / mg) og de ulike konsentrasjoner av MLA på hvert tidspunkt. Nivåer er rapportert som betyr + / - SEM, * p <0,001.
- Smak spor av DLM responser ved 100 Hz stimulering. Top spor viser svarene av muskelen før MLA injeksjon på GF stimulering i hjernen. Legg merke til at muskelen er i stand til å svare til hver stimulus en-til-en på 100 Hz. Midt spor viser svarene på DLM etter MLA injeksjon (0,12 ng / mg). Legg merke til at muskelen ikke er i stand til å svare til hver stimulus en-til-en på 100 Hz. (Stjerner). Bunnekko viser svarene fra DLM av samme forberedelse (0,12 ng / mg) ved direkte stimulering av motoriske nerveceller i thorax. Fordi DLM svarer en-til-en på 100 Hz med thorax stimulering, kan svikt i svarene med hjernen stimuleringer tilskrives kolinerg PSI-DLMn forbindelse.
- En grafisk framstilling av virkningene med ulike MLA konsentrasjoner (0, 0,02, 0,04, 0,08, 0,12 ng / mg) på GF-TTM veien. Ingen signifikante effekter ble sett mellom saltvann (0 ng / mg) og sammensatte injeksjoner som helst punkt. En t-test ble utført mellom saltvann kontroller (0 ng / mg) og de ulike konsentrasjoner av MLA på hvert tidspunkt, * p <0,001.
- Smak spor av TTM responser ved 100 Hz stimulering. Top spor viser svarene av muskelen før MLA injeksjon på GF aktivering med stimulering i hjernen. Legg merke til at muskelen er i stand til å svare på alle stimuli på 100 Hz. Midt spor viser svarene på DLM etter MLA injeksjon (0,12 ng / mg). Svarene fra TTM muskel til stimulering av GF i hjernen fortsatt en-til-en. Bunnekko viser svarene fra TTM av samme forberedelse til 100 Hz stimulering av motoriske nerveceller i thorax (0,12 ng / mg).
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Den nanoinjection / elektrofysiologi bioassay presenteres her åpner for en rask screening av forbindelser i nervesystemet av bananflue. Dette er en roman in vivo teknikk som krever små mengder av et sammensatt for å lokke fram en effekt på en rekke molekylære mål i en godt karakterisert neuronal krets. Denne metoden kan brukes til å teste bioaktivitet av ulike forbindelser, fra ukjente giftstoffer til kommersielt tilgjengelige farmakologiske agenter.
Her demonstrerte vi funksjonen til analysen vår hjelp MLA, som hadde en effekt på Giant Fiber System (GFS) av bananflue (figur 5). Vi fant at det selektivt forstyrret GF til DLM pathway men ikke GF til TTM veien. Aktivisere de motoriske nervecellene direkte via thorax stimulering viste at mangelen på GF til DLM pathway ikke skyldes en dysfunksjon i den nevromuskulære krysset (NMJ), men det var i overensstemmelse med antagonistiske effekt av MLA på Dα7 nAChR subtyper presentere på PSI-DLMn synapse (figur 1). Selv om GF til TTMn forbindelse ble vist å være kolinerge, er det ukjent hvorvidt Dα7 nAChR subenheter er til stede på dette synapse. Videre den genetiske fravær av kolin acetyltransferase (Cha) genet eller Dα7 nAChR undertype (Dα7) genet ikke forstyrre funksjonen til GF-TTMn forbindelse på grunn av samtidig tilstedeværelse av en elektrisk veikryss 5,6,17,19, 20, noe som gjør veien usannsynlig å bli påvirket av MLA.
Etter sammensatte injeksjon, bør løsningen umiddelbart dypp hele nervesystemet av dyret på grunn av sin åpne sirkulasjonssystemet 21. Hvis skikkelig injisert, når det sammensatte vanligvis thorax og buk i løpet av sekunder, men en homogen dispersjon kan ta opptil ett minutt. Men hvis det sammensatte ikke injiseres ordentlig inn i hemolymph (dvs. injisere mikropipette for deep går inn i hjernevevet) deretter langsommere spredning over hele dyret er observert. Selv fargestoff kan brukes til å praktisere en riktig injeksjonsteknikk som vist i videoen, er det ikke anbefalt å co-injisere blå konditorfarge med en sammensatt for å bli testet som det kan endre egenskapene til den sammensatte og dermed dens bioaktivitet. I tillegg, siden de fleste løsningene som brukes som løsemiddel er klare i fargen (saltvann, DMSO, etc), er det vanskelig å se hvorvidt forbindelsen ble kastet ut fra kanylen. Derfor, når oppløsende en bestemt sammensetning er det viktig å sikre at den går helt inn i løsningen, ellers uoppløste partikler vil raskt tette injeksjonen spissen, hindre utstøting fra noen av væske. Videre, selv om compound spredning kan skje umiddelbart hele hemolymph, nå de målene i det sentrale nervesystemet, så vel som nådde sin maksimale dosering, kan ta lenger basert på sammensatte kjemiske egenskaper, slik som størrelse og polarity, og dens evne til å gjennomsyre flua blod hjerne barrieren. 22 Derfor er det viktig å overvåke mulige effekter av ukjente forbindelser i flere minutter etter injeksjon, fordi ulike stoffer kan ha variasjon i tider med innsettende effekt, noe som kan øke over tid i noen tilfeller . Sterke og umiddelbare virkninger av det sammensatte som fullstendig blokkerer funksjonen av nevroner kan allerede ses med de utløste responser ved 1 Hz, mens stimulering av GFS ved høyere frekvenser (100 Hz) brukes til å oppdage mer subtile effekter på grunn av lavere dosering eller potens av et sammensatt. Hvis ingen effekter er observert etter sammensatte injeksjon kan det være på grunn av enten liten stoffet dosering eller det faktum at det sammensatte spesifikke molekylære målet ikke er til stede i GFS.
Dessuten, når du bruker bioassay presenteres her som et screeningverktøy for romanen forbindelser (for eksempel conotoxins) er det viktig å merke seg at analysen er begrenset til de molekylære mål som finnes iGFS på flua. Selv om analysen i seg selv ikke tillater å finne de faktiske molekylære mål av det sammensatte injisert, skulle tillate det for innsnevring ned av potensielle mål innenfor GFS. Ytterligere tester, for eksempel plaster klemme på nervecellene eller muskler eller genetiske interaksjonsstudier med Drosophila melanogaster mutanter, kan gjøres for å fastslå den konkrete målet for disse forbindelsene. Til slutt ble det presentert innspillingen protokoll laget for å oppdage antagonistiske effekter på funksjon GFS. Imidlertid kan innspillingen protokollen enkelt justeres for å overvåke for agonistiske effekter ved passivt å overvåke for svar indusert av det sammensatte heller enn å teste om GFS er ikke i stand til å svare pålitelig når kretsen stimuleres av eksperimentator.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Ingen interessekonflikter erklært.
Acknowledgments
Vi ønsker å takke medlemmene av Mari lab og Godenschwege lab, særlig Aline Yonezawa, for kommentarer og hjelp med denne protokollen. Dette arbeidet ble finansiert av National Institute for nevrologiske lidelser og Stroke stipend R21NS06637 til FM og TAG; AB ble finansiert av National Science Foundation award nummer 082925, URM: Integrative Biology for fremtidige forskere.
Materials
| Name | Company | Catalog Number | Comments |
| Recording glass electrodes: borosilicate glass capillaries | World Precision Instruments, Inc. | 1B100F-4 | 1.0mm OD, 0.58mm ID |
| Stimulator | Grass Technologies | Model S48 | |
| Amplifier | Getting Instruments, Inc. | Model 5A | |
| Data acquisition Software: Digidata | Molecular Devices | Model 1440A | |
| Data collection software: pCLAMP | Molecular Devices | Version 10 | |
| Stereomicroscope with fiber optic microscope ring illuminator | AmScope | SM-4T Model HL250-AR | |
| Dissecting scope for mounting | AmScope | SM-2TZ | |
| Kite Manual Micromanipulator & Tilting Base | World Precision Instruments, Inc. | Model # M3301 Kite: Model # KITE-M3-L | |
| Drosophila melanogaster Wild 10E genotype (wild type strain) | Bloomington Stock center | Stock # 3892 | |
| Vertical pipette puller | David Kopf Instruments | Model 700c | |
| Injection glass micropipettes: Borosilicate glass capillaries | World Precision Instruments, Inc. | Catalogue # 4878 | 1.14mm OD, 0.5mm ID |
| Silicon oil | Fisher Scientific | Catalogue # S159-500 | |
| Beveler | Sutter Instrument Co. | K.T. Brown Type Model # BV-10 | |
| Nanoliter2000 | World Precision Instruments, Inc. | Catalogue # B203XVY | |
| Blue food coloring | McCormick & Co. | N/A | Ingredients: Water, Propylene Glycol, FD&C Blue 1, and 0.1% Propylparaben (preservative). |
| Methyllycaconitine citrate (MLA) | Tocris Bioscience | Catalogue # 1029 | |
| Plastic wax sticks | Hygenic Corporation (Akron Ohio USA) |
References
- Koehn, F. E., Carter, G. T. The evolving role of natural products in drug discovery. Nat. Rev. Drug Discov. 4, 206-220 (2005).
- Miljanich, G. P. Ziconotide: neuronal calcium channel blocker for treating severe chronic pain. Curr. Med. Chem. 11, 3029-3040 (2004).
- Layer, R. T., Wagstaff, J. D., White, H. S. Conantokins: peptide antagonists of NMDA receptors. Curr. Med. Chem. 11, 3073-3084 (2004).
- Lewis, R. J. Conotoxins as selective inhibitors of neuronal ion channels, receptors and transporters. IUBMB Life. 56, 89-93 (2004).
- Allen, M. J., Godenschwege, T. A., Tanouye, M. A., Phelan, P. Making an escape: development and function of the Drosophila giant fibre system. Semin. Cell Dev. Biol. 17, 31-41 (2006).
- Fayyazuddin, A., Zaheer, M. A., Hiesinger, P. R., Bellen, H. J. The nicotinic acetylcholine receptor Dalpha7 is required for an escape behavior in Drosophila. PLoS biology. 4, e63 (2006).
- Jan, L. Y., Jan, Y. N. L-glutamate as an excitatory transmitter at the Drosophila larval neuromuscular junction. The Journal of physiology. 262, 215-236 (1976).
- Usherwood, P. N., Machili, P., Leaf, G. L-Glutamate at insect excitatory nerve-muscle synapses. Nature. 219, 1169-1172 (1968).
- Marrus, S. B., Portman, S. L., Allen, M. J., Moffat, K. G., DiAntonio, A. Differential localization of glutamate receptor subunits at the Drosophila neuromuscular junction. The Journal of neuroscience : the official journal of the Society for Neuroscience. 24, 1406-1415 (2004).
- Petersen, S. A., Fetter, R. D., Noordermeer, J. N., Goodman, C. S., DiAntonio, A. Genetic analysis of glutamate receptors in Drosophila reveals a retrograde signal regulating presynaptic transmitter release. Neuron. 19, 1237-1248 (1997).
- Qin, G. Four different subunits are essential for expressing the synaptic glutamate receptor at neuromuscular junctions of Drosophila. The Journal of neuroscience : the official journal of the Society for Neuroscience. 25, 3209-3218 (2005).
- Schuster, C. M. Molecular cloning of an invertebrate glutamate receptor subunit expressed in Drosophila muscle. Science. 254, 112-114 (1991).
- Tanouye, M. A., Wyman, R. J. Motor outputs of giant nerve fiber in Drosophila. Journal of. 44, 405-421 (1980).
- Augustin, H., Allen, M. J., Partridge, L. Electrophysiological Recordings from the Giant Fiber Pathway of D. melanogaster. J. Vis. Exp. (47), e2412 (2011).
- Allen, M. J., Godenschwege, T. Drosophila Neurobiology. Zhang, B., Freeman, M. R., Waddell, S. , Cold Spring Harbor Laboratory Press. 215-224 (2010).
- Blagburn, J. M., Alexopoulos, H., Davies, J. A., Bacon, J. P. Null mutation in shaking-B eliminates electrical, but not chemical, synapses in the Drosophila giant fiber system: a structural study. J. Comp. Neurol. 404, 449-458 (1999).
- Thomas, J. B., Wyman, R. J. Mutations altering synaptic connectivity between identified neurons in Drosophila. J. Neurosci. 4, 530-538 (1984).
- Baird, D. H., Schalet, A. P., Wyman, R. J. The Passover locus in Drosophila melanogaster: complex complementation and different effects on the giant fiber neural pathway. Genetics. 126, 1045-1059 (1990).
- Gorczyca, M., Hall, J. C. Identification of a cholinergic synapse in the giant fiber pathway of Drosophila using conditional mutations of acetylcholine synthesis. J. Neurogenet. 1, 289-313 (1984).
- Allen, M. J., Murphey, R. K. The chemical component of the mixed GF-TTMn synapse in Drosophila melanogaster uses acetylcholine as its neurotransmitter. The European journal of neuroscience. 26, 439-445 (2007).
- Mejia, M. A novel approach for in vivo screening of toxins using the Drosophila Giant Fiber circuit. Toxicon. 56, 1398-1407 (2010).
- Stork, T. Organization and function of the blood-brain barrier in Drosophila. J. Neurosci. 28, 587-597 (2008).

{kind=link}