The trace recordings of action potentials from Ae. aegypti gustatory sensilla (Figure 1) demonstrate the effectiveness of direct stimulation with a range of chemicals. This technique can be used to quantify responses to any stimulating chemical by counting spikes of a given amplitude and duration over a reasonable time range (generally less than 500 ms). Trace recordings must be readily reproducible under a given set of experimental conditions. Otherwise, the observed physiological responses may represent uncommon phenotypes among individuals of the test species.
Raw RNA-seq data requires filtration and accurate mapping before being presented as rounded RPKM (the number of mapped reads per kilobase length of transcript per million reads mapped) or FPKM (the number of read fragments per kilobase length of transcript per million reads mapped) numerical values (Figure 2). The distinction between these two RNA-seq normalization methods has been described in two seminal works10,11. One advantage of total mRNA sequencing is the ability to detect the expression of large gene families simultaneously. The visual presentation of the expression of many genes at once allows for a unique understanding of the molecular toolset for a given tissue. Comparisons between sexes, developmental stages or tissue types can be made rapidly.
Biological replicates for RNA-seq datasets may be limited by cost or difficulty of tissue collection. qRT-PCR offers the ability to validate small subsets of total gene expression at less cost and requires less source RNA. Side by side data comparisons (Figure 3A) allow for greater confidence in RNA-seq results, as the two methods rely on different chemistries for gene abundance estimation. Statistical equivalence functions (Figure 3B) demonstrate the limits of certainty in each case.
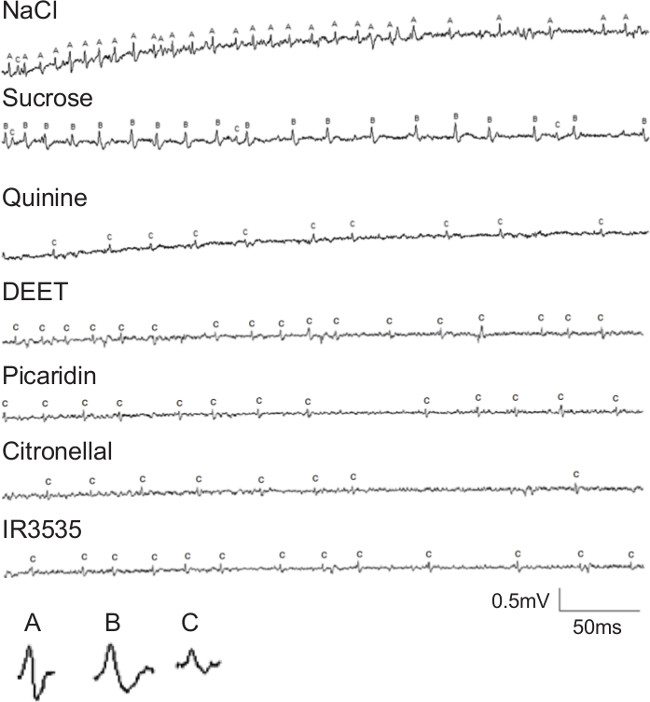
Figure 1: Electrophysiological recordings from a medium-sized sensillum on the labellum of Ae. aegypti females, modified from cited work12. Traces show responses to 100 mM NaCl, 100 mM sucrose, 1 mM quinine, 0.8% DEET, 0.8% Picaridin, 0.8% Citronellal and 0.8% IR3535. The three spikes with differing amplitudes or shapes are labeled a, b or c. These three spikes correspond with three sensory neuron subtypes with differing sensitivities: salt, sugar, and bitter, respectively.
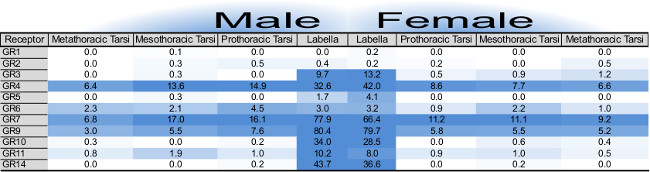
Figure 2: Table of gustatory receptor gene expression as determined by RNA-seq for 8 tissue samples, modified from cited work13. Individual cells report RPKM values for each gustatory gene annotation, male tissues (left) and female tissues (right). Numerical values are rounded to the nearest tenth. Heat-map color intensity is capped at highest RPKM and the midpoint is scaled for each gene family individually. There is only one gene family shown in this case. Please click here to view a larger version of this figure.
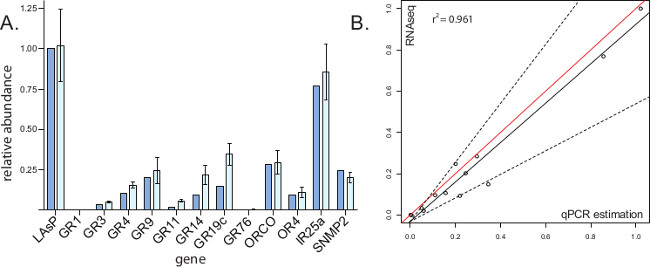
Figure 3: Comparison of qRT-PCR data and RNA-seq data, modified from cited work13. (A) Each chemoreception gene of the horizontal axis is represented by two data columns. The left column represents relative expression as determined by RNA-seq; RPKM values of chemoreception genes were divided by that of housekeeping gene AaegLAsP to generate numerical proportions in this tissue. The right column represents relative expression as determined by qRT-PCR. First, biological replicate Ct values for labella genes were normalized using AaegLAsP as a standard. Second, replicate Ct values for each gene were used to calculate relative expression values (Etarget-(Ct[target]-Ct[reference])). The error bars indicate standard deviation of individually calculated relative expression values. (B) RNA-seq and qRT-PCR datasets were compared in a separate analysis. The vertical axes represent relative gene abundance as demonstrated by RNA-seq data, with a value of 1 being attributed to housekeeping gene AaegLAsP and all other gene expression being reported relative to this value. The horizontal axes represent relative gene abundance as predicted by qRT-PCR, with a value of 1 being attributed to housekeeping gene AaegLAsP and all other gene expression being reported relative to this value. Red lines represent exact equivalence. Dashed lines represent boundaries of the 'region of similarity' for slope, calculated by the statistical equivalence algorithm9. Solid black lines represent the least-squares regression of qRT-PCR equivalence to RNA-seq quantification for the 12 target genes and AaegLAsP housekeeping gene, each represented by circle data point. Please click here to view a larger version of this figure.
